[1]
BALAT M. Status of fossil energy resources: a global perspective[J]. Energy Sources, Part B: Economics, Planning, and Policy, 2007, 2(1): 31-47.
[本文引用: 1]
[2]
ZOU C N, ZHAO Q, ZHANG G S, et al. Energy revolution: from a fossil energy era to a new energy era[J]. Natural Gas Industry B, 2016, 3(1): 1-11.
[3]
BARNES D F, FLOOR W M. Rural energy in developing countries: a challenge for economic development[J]. Annual Review of Energy and the Environment, 1996, 21(1): 497-530.
[4]
KAMPA M, CASTANAS E. Human health effects of air pollution[J]. Environmental Pollution, 2008, 151(2): 362-367.
[5]
NEWBERY D M. Acid rain[J]. Economic Policy, 1990, 5(11): 297-346.
[6]
AGGARWAL A, KUMARI R, MEHLA N, et al. Depletion of the ozone layer and its consequences: a review[J]. American Journal of Plant Sciences, 2013, 4(10): 1990.
[7]
TITUS J G. Greenhouse effect, sea level rise, and coastal zone management[J]. Coastal Management, 1986, 14(3): 147-171.
[8]
周广胜. 气候变化对中国农业生产影响研究展望[J]. 气象与环境科学, 2015, 38(1): 80-94.
ZHOU G S. Research prospect on impact of climate change on agricultural production in China[J]. Meteorological and Environmental Sciences, 2015, 38(1): 80-94.
[9]
CAMPOS A, FERNANDES C. The geopolitics of energy[J]. Geopolitics of Energy and Energy Security, 2017, 24: 23-40.
[本文引用: 1]
[10]
ILHAM N I, HASANUZZAMAN M, MAMUN M A A. World energy policies[M]// Energy for Sustainable Development. Academic Press, 2020: 179-198.
[本文引用: 1]
[11]
PAZHERI F R, OTHMAN M F, MALIK N H. A review on global renewable electricity scenario[J]. Renewable and Sustainable Energy Reviews, 2014, 31: 835-845.
[本文引用: 1]
[12]
KUMAR S, KATORIA D, SEHGAL D. Environment impact assessment of thermal power plant for sustainable development[J]. International Journal of Environmental Engineering and Management, 2013, 4(6): 567-572.
[本文引用: 1]
[13]
LAKATOS L, HEVESSY G, KOVÁCS J. Advantages and disadvantages of solar energy and wind-power utilization[J]. World Futures, 2011, 67(6): 395-408.
[本文引用: 1]
[14]
SCHERTL P, BRAUN H P. Respiratory electron transfer pathways in plant mitochondria[J]. Frontiers in Plant Science, 2014, 5: 163.
[本文引用: 1]
[15]
LEIBL W, MATHIS P. Electron transfer in photosynthesis[J]. Series on Photoconversion of Solar Energy, 2004, 2: 117.
[本文引用: 1]
[16]
MIYASAKA T, WATANABE T, FUJISHIMA A, et al. Highly efficient quantum conversion at chlorophyll a-lecithin mixed monolayer coated electrodes[J]. Nature, 1979, 277(5698): 638-640.
[本文引用: 1]
[17]
秦淑琪, 宋玉民, 王文库, 等. 水果电池的研究[J]. 甘肃农业, 2006(5): 177-177.
[本文引用: 1]
QING S Q, SONG Y M, WANG W K, et al. Research on fruit batteries[J]. Gansu Agriculture, 2006(5): 177-177.
[本文引用: 1]
[18]
GOLBERG A, RABINOWITCH H D, RUBINSKY B. Zn/Cu-vegetative batteries, bioelectrical characterizations, and primary cost analyses[J]. Journal of Renewable and Sustainable Energy, 2010, 2(3): 033103.
[本文引用: 1]
[19]
MIR L M. Therapeutic perspectives of in vivo cell electropermeabilization[J]. BioElectrochEmistry, 2001, 53(1): 1-10.
[本文引用: 1]
[20]
WEAVER J C. Electroporation of cells and tissues[J]. IEEE Transactions on Plasma Science, 2000, 28(1): 24-33.
[本文引用: 1]
[21]
KUMARA K S J, WIJESUNDERA R P, JAYASURIYA K D. Plantain pith battery powered lighting system[C]// Proceedings of the Technical Sessions, 2015, 31: 29-37.
[本文引用: 1]
[22]
KHAN K A, HASSAN L, OBAYDULLAH A K M, et al. Bioelectricity: a new approach to provide the electrical power from vegetative and fruits at off-grid region[J]. Microsystem Technologies, 2020, 26(10): 3161-3172.
[本文引用: 1]
[23]
TALAI S M, SIAGI Z O, KIMUTAI S K, et al. Comparative energy generation of Irish-potato, tomato and pineapple ZN/CU vegetative batteries[J]. Research Journal of Applied Sciences, Engineering and Technology, 2014, 8(1): 9-19.
[本文引用: 1]
[24]
CHANDNA R, AZOOZ M M, AHMAD P. Recent advances of metabolomics to reveal plant response during salt stress[J]. Salt Stress in Plants, 2013: 1-14.
[本文引用: 1]
[25]
GAO J, FENG Y P, GUO W, et al. Nanofluidics in two-dimensional layered materials: inspirations from nature[J]. Chemical Society Reviews, 2017, 46(17): 5400-5424.
[本文引用: 1]
[26]
王亚, 王海. 利用具有离子选择性的纳米通道实现浓度梯度差发电[J]. 首都师范大学学报(自然科学版), 2018, 39(6): 24-29.
[本文引用: 1]
WANG Y, WANG H. Power generation from concentration gradient using ion-selective nanochannels[J]. Journal of Capital Normal University (Natural Science Edition), 2018, 39(6): 24-29.
[本文引用: 1]
[27]
XIN W W, ZHANG Z, HUANG X D, et al. High-performance silk-based hybrid membranes employed for osmotic energy conversion[J]. Nature Communications, 2019, 10: 3876.
[本文引用: 2]
[28]
ZHANG Z, YANG S, ZHANG P P, et al. Mechanically strong MXene/Kevlar nanofiber composite membranes as high-performance nanofluidic osmotic power generators[J]. Nature Communications, 2019, 10: 2920.
[本文引用: 2]
[29]
BOCQUET L, CHARLAIX E. Nanofluidics, from bulk to interfaces[J]. Chemical Society Reviews, 2010, 39(3): 1073-1095.
[本文引用: 1]
[30]
KIM D K, DUAN C, CHEN Y F, et al. Power generation from concentration gradient by reverse electrodialysis in ion-selective nanochannels[J]. Microfluidics and Nanofluidics, 2010, 9(6): 1215-1224.
[本文引用: 2]
[31]
GHANBARI H, ESFANDIAR A. Ion transport through graphene oxide fibers as promising candidate for blue energy harvesting[J]. Carbon, 2020, 165: 267-274.
[本文引用: 2]
[32]
ZHAO Y, WANG J, KONG X Y, et al. Robust sulfonated poly (ether ether ketone) nanochannels for high-performance osmotic energy conversion[J]. National Science Review, 2020, 7(8): 1349-1359.
[本文引用: 2]
[33]
WU C R, XIAO T L, TANG J D, et al. Biomimetic temperature-gated 2D cationic nanochannels for controllable osmotic power harvesting[J]. Nano Energy, 2020, 76: 105113.
[本文引用: 2]
[34]
XIAO K, GIUSTO P, WEN L P, et al. Nanofluidic ion transport and energy conversion through ultrathin free-standing polymeric carbon nitride membranes[J]. Angewandte Chemie International Edition, 2018, 57(32): 10123-10126.
[本文引用: 1]
[35]
JI J Z, KANG Q, ZHOU Y, et al. Osmotic power generation with positively and negatively charged 2D nanofluidic membrane pairs[J]. Advanced Functional Materials, 2017, 27(2): 1603623.
[本文引用: 1]
[36]
HOU S H, ZHANG Q R, ZHANG Z, et al. Charged porous asymmetric membrane for enhancing salinity gradient energy conversion[J]. Nano Energy, 2021, 79: 105509.
[本文引用: 1]
[37]
GAO J, GUO W, FENG D, et al. High-performance ionic diode membrane for salinity gradient power generation[J]. Journal of the American Chemical Society, 2014, 136(35): 12265-12272.
[本文引用: 1]
[38]
GOVINDJEE G, WHITMARSH J. Introduction to photosynthesis: energy conversion by plants and bacteria[M]// Photosynthesis, 1982: 1-16.
[本文引用: 1]
[39]
DE CAPRARIIS B, DE FILIPPIS P, DI BATTISTA A, et al. Exoelectrogenic activity of a green microalgae, Chlorella vulgaris , in a bio-photovoltaic cells (BPVS)[J]. Chemical Engineering Transactions, 2014, 38: 523-528.
[本文引用: 1]
[40]
SAAR K L, BOMBELLI P, LEA-SMITH D J, et al. Enhancing power density of biophotovoltaics by decoupling storage and power delivery[J]. Nature Energy, 2018, 3(1): 75-81.
[本文引用: 1]
[41]
SAWA M, FANTUZZI A, BOMBELLI P, et al. Electricity generation from digitally printed cyanobacteria[J]. Nature Communications, 2017, 8(1): 1-10.
[本文引用: 1]
[42]
BOMBELLI P, MÜLLER T, HERLING T W, et al. A high power-density, mediator-free, microfluidic biophotovoltaic device for cyanobacterial cells[J]. Advanced Energy Materials, 2015, 5(2): 1401299.
[本文引用: 1]
[43]
BINDER A. Respiration and photosynthesis in energy-transducing membranes of cyanobacteria[J]. Journal of Bioenergetics and Biomembranes, 1982, 14(5): 271-286.
[本文引用: 1]
[44]
NG F L, PHANG S M, PERIASAMY V, et al. Algal biophotovoltaic (BPV) device for generation of bioelectricity using Synechococcus elongatus (Cyanophyta)[J]. Journal of Applied Phycology, 2018, 30(6): 2981-2988.
[本文引用: 1]
[45]
ZHU H W, MENG H K, ZHANG W, et al. Development of a longevous two-species biophotovoltaics with constrained electron flow[J]. Nature Communications, 2019, 10: 4282.
[本文引用: 1]
[46]
THONG C H, PHANG S M, NG F L, et al. Effect of different irradiance levels on bioelectricity generation from algal biophotovoltaic (BPV) devices[J]. Energy Science & Engineering, 2019, 7(5): 2086-2097.
[本文引用: 1]
[47]
SERVICE R F. Is it time to shoot for the sun?[J]. Science, 2005, 309(5734): 548-551.
[本文引用: 1]
[48]
GUL M M, AHMAD K S. Bioelectrochemical systems: Sustainable bio-energy powerhouses[J]. Biosensors and Bioelectronics, 2019, 142: 111576.
[本文引用: 1]
[49]
NEILSON A H, LEWIN R A. The uptake and utilization of organic carbon by algae: An essay in comparative biochemistry[J]. Phycologia, 1974, 13(3): 227-264.
[本文引用: 1]
[50]
KIM B H, CHANG I S, GADD G M. Challenges in microbial fuel cell development and operation[J]. Applied Microbiology and Biotechnology, 2007, 76(3): 485-494.
[本文引用: 1]
[51]
SMUCKER A J M. Carbon utilization and losses by plant root systems[J]. Roots, Nutrient and Water Influx, and Plant Growth, 1984, 49: 27-46.
[本文引用: 1]
[52]
宋天顺, 晏再生, 胡颖, 等. 沉积物微生物燃料电池修复水体沉积物研究进展[J]. 现代化工, 2009, 29(11): 15-19.
[本文引用: 1]
SONG T S, YAN Z S, HU Y, et al. Progress in aquatic sediment remediation by sediment microbial fuel cell[J]. Modern Chemical Industry, 2009, 29(11): 15-19.
[本文引用: 1]
[53]
KAKU N, YONEZAWA N, KODAMA Y, et al. Plant/microbe cooperation for electricity generation in a rice paddy field[J]. Applied Microbiology and Biotechnology, 2008, 79(1): 43-49.
[本文引用: 2]
[54]
SCHAMPHELAIRE L D, BOSSCHE L V, DANG H S, et al. Microbial fuel cells generating electricity from rhizodeposits of rice plants[J]. Environmental Science & Technology, 2008, 42(8): 3053-3058.
[本文引用: 4]
[55]
STRIK D P, HAMELERS H V M, SNEL J F H, et al. Green electricity production with living plants and bacteria in a fuel cell[J]. International Journal of Energy Research, 2008, 32(9): 870-876.
[本文引用: 3]
[56]
许鹏, 许丹, 张义, 等. 植物型微生物燃料电池研究进展[J]. 工业安全与环保, 2014(9): 33-35.
[本文引用: 1]
XU P, XU D, ZHANG Y, et al. Research progress of plant type microbial fuel cell[J]. Industrial Safety and Environmental Protection, 2014(9): 33-35.
[本文引用: 1]
[57]
CASTRESANA P A, MARTINEZ S M, FREEMAN E, et al. Electricity generation from moss with light-driven microbial fuel cells[J]. Electrochimica Acta, 2019, 298: 934-942.
[本文引用: 1]
[58]
IEROPOULOS I, WINFIELD J, GREENMAN J. Effects of flow-rate, inoculum and time on the internal resistance of microbial fuel cells[J]. Bioresource Technology, 2010, 101(10): 3520-3525.
[本文引用: 1]
[59]
SALINAS-JUÁREZ M G, ROQUERO P, DURÁN-DOMÍNGUEZ-DE-BAZÚA M D C. Plant and microorganisms support media for electricity generation in biological fuel cells with living hydrophytes[J]. Bioelectrochemistry, 2016, 112: 145-152.
[本文引用: 2]
[60]
TIMMERS R A, STRIK D P, HAMELERS H V M, et al. Long-term performance of a plant microbial fuel cell with Spartina anglica [J]. Applied Microbiology and Biotechnology, 2010, 86(3): 973-981.
[本文引用: 3]
[61]
LU Z H, YIN D, CHEN P, et al. Power-generating trees: Direct bioelectricity production from plants with microbial fuel cells[J]. Applied Energy, 2020, 268: 115040.
[本文引用: 1]
[62]
POWELL R J, WHITE R, HILL R T. Merging metabolism and power: development of a novel photobioelectric device driven by photosynthesis and respiration[J]. Plos One, 2014, 9(1): e86518.
[本文引用: 1]
[63]
张廷滔, 张礼霞, 高平, 等. 混合菌群生物燃料电池的产电机理与特性[J]. 应用与环境生物学报, 2012(3): 465-470.
[本文引用: 1]
ZHANG T T, ZHANG L X, GAO P, et al. Mechanism and characteristics of electricity generation in microbial fuel cells catalyzed by mixed culture[J]. Chinese Journal of Applied and Environmental Biology, 2012(3): 465-470.
[本文引用: 1]
[64]
LIU Y, DING M Z, LING W, et al. A three-species microbial consortium for power generation[J]. Energy & Environmental Science, 2017, 10(7): 1600-1609.
[本文引用: 1]
[65]
HELDER M, STRIK D, HAMELERS H V M, et al. Concurrent bio-electricity and biomass production in three plant-microbial fuel cells using Spartina anglica , Arundinella anomala and Arundo donax [J]. Bioresource Technology, 2010, 101(10): 3541-3547.
[本文引用: 1]
[66]
付哲平, 李子健. 海泥细菌电池技术原理与特点及其应用前景分析研究——一种新型海洋可再生能源技术[J]. 海洋开发与管理, 2017, 34(3): 67-71.
[本文引用: 1]
FU Z P, LI Z J. Principle and characteristic of technology and its application prospect of marine sediment microbial fuel cell[J]. Ocean Development and Management, 2017, 34(3): 67-71.
[本文引用: 1]
[67]
LOGAN B E. Microbial fuel cells[M]. John Wiley & Sons, 2008.
[本文引用: 1]
[68]
PRAKASH O, MUNGRAY A, KAILASA S K, et al. Comparison of different electrode materials and modification for power enhancement in benthic microbial fuel cells (BMFCs)[J]. Process Safety and Environmental Protection, 2018, 117: 11-21.
[本文引用: 1]
[69]
FU Y B, XU Q, ZAI X R, et al. Low electrical potential anode modified with Fe/ferric oxide and its application in marine benthic microbial fuel cell with higher voltage and power output[J]. Applied Surface Science, 2014, 289: 472-477.
[本文引用: 1]
[70]
FU Y B, LIU Z H, SU G, et al. Modified carbon anode by MWCNTs/PANI used in marine sediment microbial fuel cell and its electrochemical performance[J]. Fuel Cells, 2016, 16(3): 377-383.
[本文引用: 1]
[71]
STRIK D P, TIMMERS R A, HELDER M, et al. Microbial solar cells: Applying photosynthetic and electrochemically active organisms[J]. Trends in Biotechnology, 2011, 29(1): 41-49.
[本文引用: 1]
[72]
MOHAN S V, MOHANAKRISHNA G, CHIRANJEEVI P. Sustainable power generation from floating macrophytes based ecological microenvironment through embedded fuel cells along with simultaneous wastewater treatment[J]. Bioresource Technology, 2011, 102(14): 7036-7042.
[本文引用: 1]
[73]
毛慧, 钱伟东. 水稻集中育供秧的现状与探索[J]. 农业与技术, 2018, 38(15): 167-168.
[本文引用: 1]
MAO H, QIAN W D. Current situation and exploration of centralized breeding and supply of rice seedlings[J]. Agriculture and Technology, 2018, 38(15): 167-168.
[本文引用: 1]
[74]
ORMEROD R M. Solid oxide fuel cells[J]. Chemical Society Reviews, 2003, 32(1): 17-28.
[本文引用: 1]
[75]
AELTERMAN P, RABAEY K, CLAUWAERT P, et al. Microbial fuel cells for wastewater treatment[J]. Water Science and Technology, 2006, 54(8): 9-15.
[本文引用: 1]
[76]
MAAMER B, BOUGHAMOURA A, FATH EL-BAB A M R, et al. A review on design improvements and techniques for mechanical energy harvesting using piezoelectric and electromagnetic schemes[J]. Energy Conversion and Management, 2019, 199: 111973.
[本文引用: 1]
[77]
ZI Y L, GUO H Y, WEN Z, et al. Harvesting low-frequency (<5 Hz) irregular mechanical energy: A possible killer application of triboelectric nanogenerator[J]. ACS Nano, 2016, 10(4): 4797-4805.
[本文引用: 1]
[78]
KNIGHT M R, SMITH S M, TREWAVAS A J. Wind-induced plant motion immediately increases cytosolic calcium[J]. Proceedings of the National Academy of Sciences, 1992, 89(11): 4967-4971.
[本文引用: 1]
[79]
POHANKA M. The piezoelectric biosensors: Principles and applications[J]. International Journal of Electrochemical Science, 2017, 12: 496-506.
[本文引用: 1]
[80]
PAN S H, ZHANG Z N. Fundamental theories and basic principles of triboelectric effect: a review[J]. Friction, 2019, 7(1): 2-17.
[本文引用: 1]
[81]
CHEN Y D, JIE Y, WANG J, et al. Triboelectrification on natural rose petal for harvesting environmental mechanical energy[J]. Nano Energy, 2018, 50: 441-447.
[本文引用: 1]
[82]
WU H, CHEN Z F, XU G Q, et al. Fully biodegradable water droplet energy harvester based on leaves of living plants[J]. ACS Applied Materials & Interfaces, 2020, 12(50): 56060-56067.
[本文引用: 1]
[83]
JIE Y, JIA X T, ZOU J D, et al. Natural leaf made triboelectric nanogenerator for harvesting environmental mechanical energy[J]. Advanced Energy Materials, 2018, 8(12): 1703133.
[本文引用: 1]
[84]
MEDER F, THIELEN M, MONDINI A, et al. Living plant-hybrid generators for multidirectional wind energy conversion[J]. Energy Technology, 2020, 8(7): 2000236.
[本文引用: 1]
[85]
YAN J, LIU M, JEONG Y G, et al. Performance enhancements in poly (vinylidene fluoride)-based piezoelectric nanogenerators for efficient energy harvesting[J]. Nano Energy, 2019, 56: 662-692.
[本文引用: 1]
[86]
SLABOV V, KOPYL S, DOS SANTOS M P S, et al. Natural and eco-friendly materials for triboelectric energy harvesting[J]. Nano-Micro Letters, 2020, 12(1): 42.
[本文引用: 1]
[87]
刘强, 许鑫华, 任光雷, 等. 酶生物燃料电池[J]. 化学进展, 2006, 18(11): 1530.
[本文引用: 1]
LIU Q, XU X H, REN G L, et al. Enzymatic biofuel cells[J]. Progress in Chemistry, 2006, 18(11): 1530.
[本文引用: 1]
1
... 随着传统化石能源的过度消耗及所产生的环境污染日渐严峻,能源问题俨然已经成为影响一个国家和地区可持续发展的重大战略问题[1 -9 ] .近年来,世界各国积极制定新能源发展战略,希望通过合理利用常规能源和研发新型清洁能源,实现能源结构的转型升级,推动经济社会稳定发展、生态环境不断改善[10 ] .能源的最主要用途之一是生产生活供电[11 ] .传统的发电技术是火力发电,采用石油、煤炭和天然气等燃料燃烧产生的热能来加热水,使水变成高温、高压的水蒸气,然后由水蒸气推动发电机来发电[12 ] .尽管具有投资少、见效快的特点,但其对环境污染严重,难以满足可持续绿色发展需求.利用清洁能源实现发电的技术,如风力发电和太阳能发电,普遍具有节能环保的特点,但这些发电技术设备造价高、地域依赖性强且易受气候影响,也有待改进[13 ] .未来社会发展对电能获取的需求可归纳为“安全高效、可持续、低成本”,这也应是对于能源发展不变的追求. ...
1
... 随着传统化石能源的过度消耗及所产生的环境污染日渐严峻,能源问题俨然已经成为影响一个国家和地区可持续发展的重大战略问题[1 -9 ] .近年来,世界各国积极制定新能源发展战略,希望通过合理利用常规能源和研发新型清洁能源,实现能源结构的转型升级,推动经济社会稳定发展、生态环境不断改善[10 ] .能源的最主要用途之一是生产生活供电[11 ] .传统的发电技术是火力发电,采用石油、煤炭和天然气等燃料燃烧产生的热能来加热水,使水变成高温、高压的水蒸气,然后由水蒸气推动发电机来发电[12 ] .尽管具有投资少、见效快的特点,但其对环境污染严重,难以满足可持续绿色发展需求.利用清洁能源实现发电的技术,如风力发电和太阳能发电,普遍具有节能环保的特点,但这些发电技术设备造价高、地域依赖性强且易受气候影响,也有待改进[13 ] .未来社会发展对电能获取的需求可归纳为“安全高效、可持续、低成本”,这也应是对于能源发展不变的追求. ...
1
... 随着传统化石能源的过度消耗及所产生的环境污染日渐严峻,能源问题俨然已经成为影响一个国家和地区可持续发展的重大战略问题[1 -9 ] .近年来,世界各国积极制定新能源发展战略,希望通过合理利用常规能源和研发新型清洁能源,实现能源结构的转型升级,推动经济社会稳定发展、生态环境不断改善[10 ] .能源的最主要用途之一是生产生活供电[11 ] .传统的发电技术是火力发电,采用石油、煤炭和天然气等燃料燃烧产生的热能来加热水,使水变成高温、高压的水蒸气,然后由水蒸气推动发电机来发电[12 ] .尽管具有投资少、见效快的特点,但其对环境污染严重,难以满足可持续绿色发展需求.利用清洁能源实现发电的技术,如风力发电和太阳能发电,普遍具有节能环保的特点,但这些发电技术设备造价高、地域依赖性强且易受气候影响,也有待改进[13 ] .未来社会发展对电能获取的需求可归纳为“安全高效、可持续、低成本”,这也应是对于能源发展不变的追求. ...
1
... 随着传统化石能源的过度消耗及所产生的环境污染日渐严峻,能源问题俨然已经成为影响一个国家和地区可持续发展的重大战略问题[1 -9 ] .近年来,世界各国积极制定新能源发展战略,希望通过合理利用常规能源和研发新型清洁能源,实现能源结构的转型升级,推动经济社会稳定发展、生态环境不断改善[10 ] .能源的最主要用途之一是生产生活供电[11 ] .传统的发电技术是火力发电,采用石油、煤炭和天然气等燃料燃烧产生的热能来加热水,使水变成高温、高压的水蒸气,然后由水蒸气推动发电机来发电[12 ] .尽管具有投资少、见效快的特点,但其对环境污染严重,难以满足可持续绿色发展需求.利用清洁能源实现发电的技术,如风力发电和太阳能发电,普遍具有节能环保的特点,但这些发电技术设备造价高、地域依赖性强且易受气候影响,也有待改进[13 ] .未来社会发展对电能获取的需求可归纳为“安全高效、可持续、低成本”,这也应是对于能源发展不变的追求. ...
1
... 随着传统化石能源的过度消耗及所产生的环境污染日渐严峻,能源问题俨然已经成为影响一个国家和地区可持续发展的重大战略问题[1 -9 ] .近年来,世界各国积极制定新能源发展战略,希望通过合理利用常规能源和研发新型清洁能源,实现能源结构的转型升级,推动经济社会稳定发展、生态环境不断改善[10 ] .能源的最主要用途之一是生产生活供电[11 ] .传统的发电技术是火力发电,采用石油、煤炭和天然气等燃料燃烧产生的热能来加热水,使水变成高温、高压的水蒸气,然后由水蒸气推动发电机来发电[12 ] .尽管具有投资少、见效快的特点,但其对环境污染严重,难以满足可持续绿色发展需求.利用清洁能源实现发电的技术,如风力发电和太阳能发电,普遍具有节能环保的特点,但这些发电技术设备造价高、地域依赖性强且易受气候影响,也有待改进[13 ] .未来社会发展对电能获取的需求可归纳为“安全高效、可持续、低成本”,这也应是对于能源发展不变的追求. ...
1
... 随着传统化石能源的过度消耗及所产生的环境污染日渐严峻,能源问题俨然已经成为影响一个国家和地区可持续发展的重大战略问题[1 -9 ] .近年来,世界各国积极制定新能源发展战略,希望通过合理利用常规能源和研发新型清洁能源,实现能源结构的转型升级,推动经济社会稳定发展、生态环境不断改善[10 ] .能源的最主要用途之一是生产生活供电[11 ] .传统的发电技术是火力发电,采用石油、煤炭和天然气等燃料燃烧产生的热能来加热水,使水变成高温、高压的水蒸气,然后由水蒸气推动发电机来发电[12 ] .尽管具有投资少、见效快的特点,但其对环境污染严重,难以满足可持续绿色发展需求.利用清洁能源实现发电的技术,如风力发电和太阳能发电,普遍具有节能环保的特点,但这些发电技术设备造价高、地域依赖性强且易受气候影响,也有待改进[13 ] .未来社会发展对电能获取的需求可归纳为“安全高效、可持续、低成本”,这也应是对于能源发展不变的追求. ...
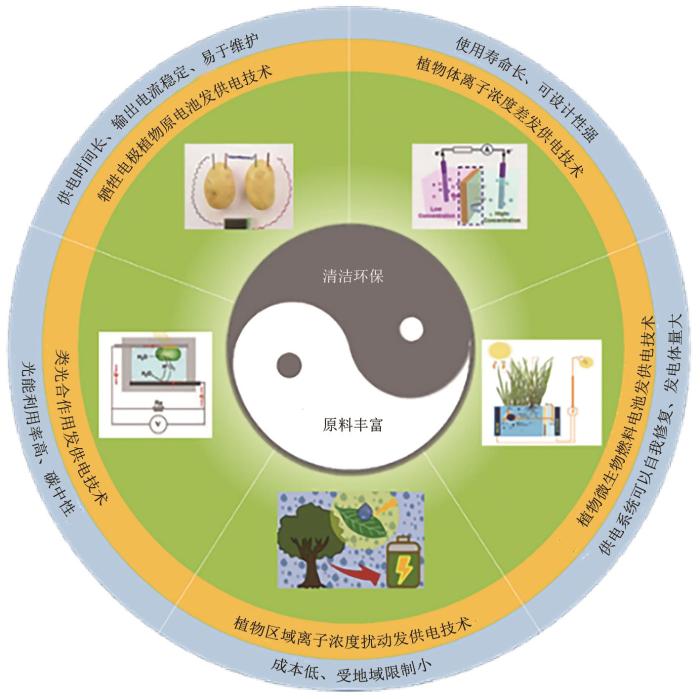
1
... 科技飞速发展拓展了电能的获取途径,植物发供电技术就是一种从植物中获取电能的新方法.该技术以植物作为发供电主体,通过电化学手段、植物体的生理过程和外界环境的干扰等,有效地将自然界中取之不尽的光能、机械能和生物质能等能量转化为电能,并且几乎不会对环境造成污染.电在植物体内普遍存在,组成植物体的每个细胞都是一台微型发电机,植物体内许多重要的生命活动都伴随着电子的产生和传递,如呼吸作用和光合作用等[14 -15 ] .在20世纪,英国的钟表匠托尼·埃希尔就发现了植物本身可以产生电流.他用铜线和锌线分别插入柠檬,令其与小型钟表上的电动机与电路相连接,使小型钟表工作了长达5个月之久.此外,日本研究人员为了验证叶绿素发电的可行性,将从菠菜中提取到的叶绿素与卵磷脂混合,把混合物涂在透明的氧化锡结晶片上,用它作为正极安置在“透明电池”中,在太阳光的照射下即产生电流,转换效率可达普通太阳能电池的3倍以上[16 ] .但是叶绿素与卵磷脂离开植物后很容易分解,失去吸收太阳能的作用,所以这款电池不能持久发电.为此,英国剑桥的研究人员直接利用活体植物进行发电,研究表明在天气晴朗情况下,一盆直径1 m的蕨类植物每天可以产生将近1度电.利用植物与附近土壤pH的差异也可以获得电流输出,荷兰瓦赫宁根大学研究人员成立的Plant-e公司基于此原理从生长的植物中不断获取电力,在夜晚点亮城市LED街灯. ...
1
... 科技飞速发展拓展了电能的获取途径,植物发供电技术就是一种从植物中获取电能的新方法.该技术以植物作为发供电主体,通过电化学手段、植物体的生理过程和外界环境的干扰等,有效地将自然界中取之不尽的光能、机械能和生物质能等能量转化为电能,并且几乎不会对环境造成污染.电在植物体内普遍存在,组成植物体的每个细胞都是一台微型发电机,植物体内许多重要的生命活动都伴随着电子的产生和传递,如呼吸作用和光合作用等[14 -15 ] .在20世纪,英国的钟表匠托尼·埃希尔就发现了植物本身可以产生电流.他用铜线和锌线分别插入柠檬,令其与小型钟表上的电动机与电路相连接,使小型钟表工作了长达5个月之久.此外,日本研究人员为了验证叶绿素发电的可行性,将从菠菜中提取到的叶绿素与卵磷脂混合,把混合物涂在透明的氧化锡结晶片上,用它作为正极安置在“透明电池”中,在太阳光的照射下即产生电流,转换效率可达普通太阳能电池的3倍以上[16 ] .但是叶绿素与卵磷脂离开植物后很容易分解,失去吸收太阳能的作用,所以这款电池不能持久发电.为此,英国剑桥的研究人员直接利用活体植物进行发电,研究表明在天气晴朗情况下,一盆直径1 m的蕨类植物每天可以产生将近1度电.利用植物与附近土壤pH的差异也可以获得电流输出,荷兰瓦赫宁根大学研究人员成立的Plant-e公司基于此原理从生长的植物中不断获取电力,在夜晚点亮城市LED街灯. ...
1
... 科技飞速发展拓展了电能的获取途径,植物发供电技术就是一种从植物中获取电能的新方法.该技术以植物作为发供电主体,通过电化学手段、植物体的生理过程和外界环境的干扰等,有效地将自然界中取之不尽的光能、机械能和生物质能等能量转化为电能,并且几乎不会对环境造成污染.电在植物体内普遍存在,组成植物体的每个细胞都是一台微型发电机,植物体内许多重要的生命活动都伴随着电子的产生和传递,如呼吸作用和光合作用等[14 -15 ] .在20世纪,英国的钟表匠托尼·埃希尔就发现了植物本身可以产生电流.他用铜线和锌线分别插入柠檬,令其与小型钟表上的电动机与电路相连接,使小型钟表工作了长达5个月之久.此外,日本研究人员为了验证叶绿素发电的可行性,将从菠菜中提取到的叶绿素与卵磷脂混合,把混合物涂在透明的氧化锡结晶片上,用它作为正极安置在“透明电池”中,在太阳光的照射下即产生电流,转换效率可达普通太阳能电池的3倍以上[16 ] .但是叶绿素与卵磷脂离开植物后很容易分解,失去吸收太阳能的作用,所以这款电池不能持久发电.为此,英国剑桥的研究人员直接利用活体植物进行发电,研究表明在天气晴朗情况下,一盆直径1 m的蕨类植物每天可以产生将近1度电.利用植物与附近土壤pH的差异也可以获得电流输出,荷兰瓦赫宁根大学研究人员成立的Plant-e公司基于此原理从生长的植物中不断获取电力,在夜晚点亮城市LED街灯. ...
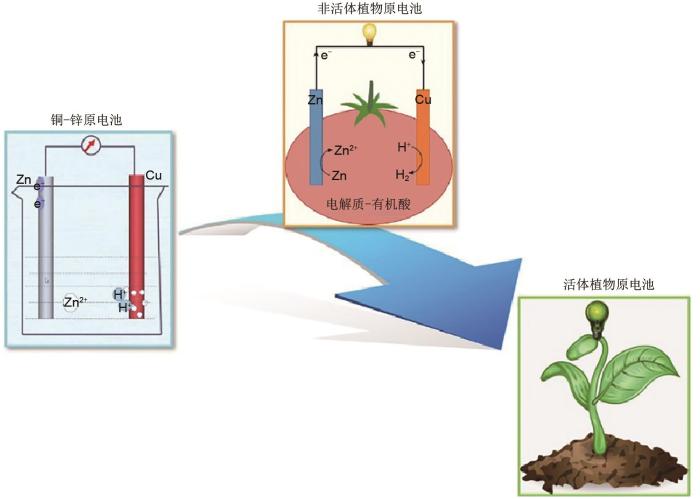
1
... 牺牲电极植物原电池发供电技术是以植物为主体,采用铜和锌作为电极、植物组织液作为电解质,组成原电池进行发供电的一种技术.其原理为:插入植物中的两种金属电极的电化学活性不同,更活泼的锌容易失去电子,置换出植物中酸性物质的氢离子(图2 ).锌电极失去部分电子后,与铜电极之间产生电位差,当用导线把两个电极连接起来,在电位差驱动下电子由锌电极通过导线流向铜电极,形成了电流[17 ] .只要植物体内参与反应物持续产生保证反应进行,就可持续输出电流. ...
1
... 牺牲电极植物原电池发供电技术是以植物为主体,采用铜和锌作为电极、植物组织液作为电解质,组成原电池进行发供电的一种技术.其原理为:插入植物中的两种金属电极的电化学活性不同,更活泼的锌容易失去电子,置换出植物中酸性物质的氢离子(图2 ).锌电极失去部分电子后,与铜电极之间产生电位差,当用导线把两个电极连接起来,在电位差驱动下电子由锌电极通过导线流向铜电极,形成了电流[17 ] .只要植物体内参与反应物持续产生保证反应进行,就可持续输出电流. ...
1
... 近年来,研究人员在该领域取得了一系列研究进展.例如,Golberg等[18 ] 设计了马铃薯铜锌原电池,分别研究了新鲜马铃薯、不可逆电穿孔处理的马铃薯和加热处理的马铃薯的放电功率、电池容量等电性能.其中,组织电穿孔(电渗透)是一种给组织细胞施加电场以增加细胞膜的渗透性,从而允许化学品、药物或DNA被引入细胞的微生物学技术,这种技术通过在目标组织内或周围放置电极,在组织内产生脉冲电场来实现[19 ] .这些电场诱导细胞膜发生可逆或不可逆的结构变化,增强药物或DNA等向细胞质的渗透能力[20 ] .研究发现,经过加热或电穿孔处理的马铃薯产生的电能是未经处理的马铃薯的10倍.该团队认为这是由于经过处理后的马铃薯细胞的细胞膜被破坏,质子的有效扩散率随着膜的破裂而增加.相反,当细胞膜完整时,质子扩散的速率降低,这可能是由于细胞内液体不能高效参与离子转运,单位体积内电解质浓度的等效降低.马铃薯原电池给低功率用电器供电的研究如图3 (a)所示.两个白色标准发光二极管(最小正向电流为2 mA,电压为1.8 V)并联在5个加热的土豆电池上,每个电池由马铃薯片夹在锌片和铜片之间组成.在这种情况下,其可以支持发光二极管连续发光3 h.Kumara等[21 ] 将车前草样品夹在平行的锌和铜电极之间制成原电池,通过改变电解质状态(车前草分别煮沸5、10、15、20、30和35 min并切碎)、电极距离(1、1.5、2和2.5 cm)和电极尺寸(5 cm×9 cm和11 cm×11 cm)等条件探索锌铜植物原电池的电性能.结果表明:车前草煮沸20 min后切碎可以有效提高锌铜植物原电池的电性能;电极间距为1.5 cm时电池电性能最佳;当电极尺寸从5 cm×9 cm增加到11 cm×11 cm时,稳态电流略有增加,且与电极尺寸的增加不成正比.该团队使用60个灰车前草锌铜原电池制作了灰车前草电池组,即并联6组,每组串联10个电池,每个电池的尺寸为5 cm×9 cm[图3 (b)].研究表明,该电池组的开路电压为3.86 V,由该电池组供电的3 W发光二极管照明系统可提供强度约为3500 Lux的恒定光照,供电时间超过600 h. ...
1
... 近年来,研究人员在该领域取得了一系列研究进展.例如,Golberg等[18 ] 设计了马铃薯铜锌原电池,分别研究了新鲜马铃薯、不可逆电穿孔处理的马铃薯和加热处理的马铃薯的放电功率、电池容量等电性能.其中,组织电穿孔(电渗透)是一种给组织细胞施加电场以增加细胞膜的渗透性,从而允许化学品、药物或DNA被引入细胞的微生物学技术,这种技术通过在目标组织内或周围放置电极,在组织内产生脉冲电场来实现[19 ] .这些电场诱导细胞膜发生可逆或不可逆的结构变化,增强药物或DNA等向细胞质的渗透能力[20 ] .研究发现,经过加热或电穿孔处理的马铃薯产生的电能是未经处理的马铃薯的10倍.该团队认为这是由于经过处理后的马铃薯细胞的细胞膜被破坏,质子的有效扩散率随着膜的破裂而增加.相反,当细胞膜完整时,质子扩散的速率降低,这可能是由于细胞内液体不能高效参与离子转运,单位体积内电解质浓度的等效降低.马铃薯原电池给低功率用电器供电的研究如图3 (a)所示.两个白色标准发光二极管(最小正向电流为2 mA,电压为1.8 V)并联在5个加热的土豆电池上,每个电池由马铃薯片夹在锌片和铜片之间组成.在这种情况下,其可以支持发光二极管连续发光3 h.Kumara等[21 ] 将车前草样品夹在平行的锌和铜电极之间制成原电池,通过改变电解质状态(车前草分别煮沸5、10、15、20、30和35 min并切碎)、电极距离(1、1.5、2和2.5 cm)和电极尺寸(5 cm×9 cm和11 cm×11 cm)等条件探索锌铜植物原电池的电性能.结果表明:车前草煮沸20 min后切碎可以有效提高锌铜植物原电池的电性能;电极间距为1.5 cm时电池电性能最佳;当电极尺寸从5 cm×9 cm增加到11 cm×11 cm时,稳态电流略有增加,且与电极尺寸的增加不成正比.该团队使用60个灰车前草锌铜原电池制作了灰车前草电池组,即并联6组,每组串联10个电池,每个电池的尺寸为5 cm×9 cm[图3 (b)].研究表明,该电池组的开路电压为3.86 V,由该电池组供电的3 W发光二极管照明系统可提供强度约为3500 Lux的恒定光照,供电时间超过600 h. ...
1
... 近年来,研究人员在该领域取得了一系列研究进展.例如,Golberg等[18 ] 设计了马铃薯铜锌原电池,分别研究了新鲜马铃薯、不可逆电穿孔处理的马铃薯和加热处理的马铃薯的放电功率、电池容量等电性能.其中,组织电穿孔(电渗透)是一种给组织细胞施加电场以增加细胞膜的渗透性,从而允许化学品、药物或DNA被引入细胞的微生物学技术,这种技术通过在目标组织内或周围放置电极,在组织内产生脉冲电场来实现[19 ] .这些电场诱导细胞膜发生可逆或不可逆的结构变化,增强药物或DNA等向细胞质的渗透能力[20 ] .研究发现,经过加热或电穿孔处理的马铃薯产生的电能是未经处理的马铃薯的10倍.该团队认为这是由于经过处理后的马铃薯细胞的细胞膜被破坏,质子的有效扩散率随着膜的破裂而增加.相反,当细胞膜完整时,质子扩散的速率降低,这可能是由于细胞内液体不能高效参与离子转运,单位体积内电解质浓度的等效降低.马铃薯原电池给低功率用电器供电的研究如图3 (a)所示.两个白色标准发光二极管(最小正向电流为2 mA,电压为1.8 V)并联在5个加热的土豆电池上,每个电池由马铃薯片夹在锌片和铜片之间组成.在这种情况下,其可以支持发光二极管连续发光3 h.Kumara等[21 ] 将车前草样品夹在平行的锌和铜电极之间制成原电池,通过改变电解质状态(车前草分别煮沸5、10、15、20、30和35 min并切碎)、电极距离(1、1.5、2和2.5 cm)和电极尺寸(5 cm×9 cm和11 cm×11 cm)等条件探索锌铜植物原电池的电性能.结果表明:车前草煮沸20 min后切碎可以有效提高锌铜植物原电池的电性能;电极间距为1.5 cm时电池电性能最佳;当电极尺寸从5 cm×9 cm增加到11 cm×11 cm时,稳态电流略有增加,且与电极尺寸的增加不成正比.该团队使用60个灰车前草锌铜原电池制作了灰车前草电池组,即并联6组,每组串联10个电池,每个电池的尺寸为5 cm×9 cm[图3 (b)].研究表明,该电池组的开路电压为3.86 V,由该电池组供电的3 W发光二极管照明系统可提供强度约为3500 Lux的恒定光照,供电时间超过600 h. ...
1
... 近年来,研究人员在该领域取得了一系列研究进展.例如,Golberg等[18 ] 设计了马铃薯铜锌原电池,分别研究了新鲜马铃薯、不可逆电穿孔处理的马铃薯和加热处理的马铃薯的放电功率、电池容量等电性能.其中,组织电穿孔(电渗透)是一种给组织细胞施加电场以增加细胞膜的渗透性,从而允许化学品、药物或DNA被引入细胞的微生物学技术,这种技术通过在目标组织内或周围放置电极,在组织内产生脉冲电场来实现[19 ] .这些电场诱导细胞膜发生可逆或不可逆的结构变化,增强药物或DNA等向细胞质的渗透能力[20 ] .研究发现,经过加热或电穿孔处理的马铃薯产生的电能是未经处理的马铃薯的10倍.该团队认为这是由于经过处理后的马铃薯细胞的细胞膜被破坏,质子的有效扩散率随着膜的破裂而增加.相反,当细胞膜完整时,质子扩散的速率降低,这可能是由于细胞内液体不能高效参与离子转运,单位体积内电解质浓度的等效降低.马铃薯原电池给低功率用电器供电的研究如图3 (a)所示.两个白色标准发光二极管(最小正向电流为2 mA,电压为1.8 V)并联在5个加热的土豆电池上,每个电池由马铃薯片夹在锌片和铜片之间组成.在这种情况下,其可以支持发光二极管连续发光3 h.Kumara等[21 ] 将车前草样品夹在平行的锌和铜电极之间制成原电池,通过改变电解质状态(车前草分别煮沸5、10、15、20、30和35 min并切碎)、电极距离(1、1.5、2和2.5 cm)和电极尺寸(5 cm×9 cm和11 cm×11 cm)等条件探索锌铜植物原电池的电性能.结果表明:车前草煮沸20 min后切碎可以有效提高锌铜植物原电池的电性能;电极间距为1.5 cm时电池电性能最佳;当电极尺寸从5 cm×9 cm增加到11 cm×11 cm时,稳态电流略有增加,且与电极尺寸的增加不成正比.该团队使用60个灰车前草锌铜原电池制作了灰车前草电池组,即并联6组,每组串联10个电池,每个电池的尺寸为5 cm×9 cm[图3 (b)].研究表明,该电池组的开路电压为3.86 V,由该电池组供电的3 W发光二极管照明系统可提供强度约为3500 Lux的恒定光照,供电时间超过600 h. ...
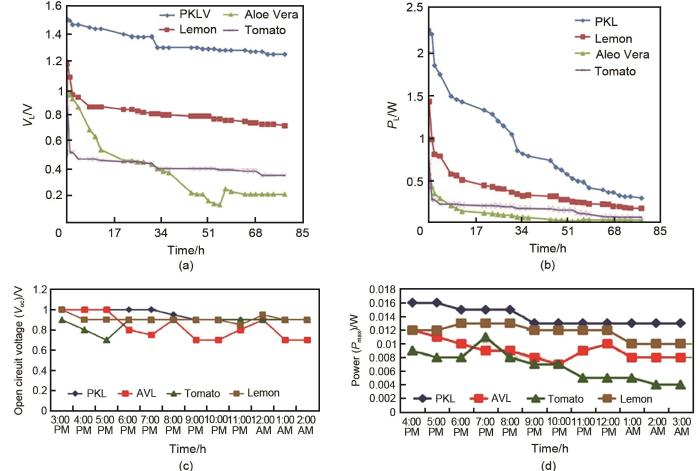
1
... 此外,研究人员探究了不同植物对铜锌原电池供电性能的影响.Khan等[22 ] 研究了苔藓叶(PKL)、芦荟、番茄和柠檬4种铜锌原电池在外加1 Ω负载电阻、锌铜电极个数比为2∶1时的电性能.从原电池的负载电压、负载功率随时间的变化可以看出,PKL电化学电池比其他3种电化学电池输出功率更高,输出电压更为稳定,3 d后仍保持在1 V以上[图4 (a)、(b)].该团队还研究了上述4种植物原电池在无负载、锌铜电极个数比为1∶1时的电性能,同样证明了PKL电化学电池优异的放电特性[图4 (c)、(d)].植物原电池温度响应的性能结果显示,在50 ℃下处理20 min可以得到最佳的电性能,说明热处理引起的细胞和组织结构的变化会影响植物原电池的电性能,改变发电量.Talai等[23 ] 分析了以马铃薯、菠萝和番茄分别作为电解质来源的铜锌原电池的发供电情况.通过对植物样品在不同条件下进行加热或电穿孔处理,确定了其电性能的变化和最佳的处理条件.第1次处理是将植物样品在不同温度(19.5~80 ℃)和不同保温时间(20~60 min)下进行水浴加热;第2次处理将样品夹在两块铝板之间,在3.12 mV的固定电压下对样品进行电穿孔,交流电的频率为2.63~100000 Hz.实验结果表明,输出电压随着加热温度和交流电频率的升高先增大后减小.对于大小为108 cm3 的样品,在60 ℃的温度下保持20 min的热处理,可产生最大开路电压:马铃薯为890 mV,菠萝为942 mV,番茄为939 mV.在最佳交流电频率(8 Hz)下,可观察到最大电流:马铃薯为0.006 mA,菠萝为0.008 mA,番茄为0.008 mA.实验还研究了植物原电池的实际供电能力.将干电池手电筒中取下的2个白色发光二极管(最小正向电流为1.44 mA,电压为3 V)和6个经过电穿孔的马铃薯电池串联,发现可以支持发光二极管连续发光1 h,直到自动断开,证明了该植物原电池供给低功率电能需求的潜力. ...
1
... 此外,研究人员探究了不同植物对铜锌原电池供电性能的影响.Khan等[22 ] 研究了苔藓叶(PKL)、芦荟、番茄和柠檬4种铜锌原电池在外加1 Ω负载电阻、锌铜电极个数比为2∶1时的电性能.从原电池的负载电压、负载功率随时间的变化可以看出,PKL电化学电池比其他3种电化学电池输出功率更高,输出电压更为稳定,3 d后仍保持在1 V以上[图4 (a)、(b)].该团队还研究了上述4种植物原电池在无负载、锌铜电极个数比为1∶1时的电性能,同样证明了PKL电化学电池优异的放电特性[图4 (c)、(d)].植物原电池温度响应的性能结果显示,在50 ℃下处理20 min可以得到最佳的电性能,说明热处理引起的细胞和组织结构的变化会影响植物原电池的电性能,改变发电量.Talai等[23 ] 分析了以马铃薯、菠萝和番茄分别作为电解质来源的铜锌原电池的发供电情况.通过对植物样品在不同条件下进行加热或电穿孔处理,确定了其电性能的变化和最佳的处理条件.第1次处理是将植物样品在不同温度(19.5~80 ℃)和不同保温时间(20~60 min)下进行水浴加热;第2次处理将样品夹在两块铝板之间,在3.12 mV的固定电压下对样品进行电穿孔,交流电的频率为2.63~100000 Hz.实验结果表明,输出电压随着加热温度和交流电频率的升高先增大后减小.对于大小为108 cm3 的样品,在60 ℃的温度下保持20 min的热处理,可产生最大开路电压:马铃薯为890 mV,菠萝为942 mV,番茄为939 mV.在最佳交流电频率(8 Hz)下,可观察到最大电流:马铃薯为0.006 mA,菠萝为0.008 mA,番茄为0.008 mA.实验还研究了植物原电池的实际供电能力.将干电池手电筒中取下的2个白色发光二极管(最小正向电流为1.44 mA,电压为3 V)和6个经过电穿孔的马铃薯电池串联,发现可以支持发光二极管连续发光1 h,直到自动断开,证明了该植物原电池供给低功率电能需求的潜力. ...
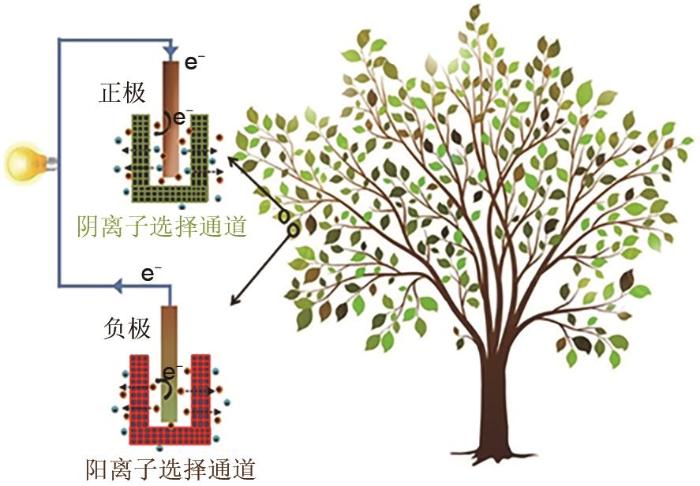
1
... 植物在维持生命活动或应对环境变化时会自动调节各部位的离子浓度,从而为生命活动提供一个相对稳定的电解质环境以确保酶的活性[24 ] .植物体离子浓度差发供电技术就是利用植物不同部位的离子浓度不同,由离子浓度差异产生电势差实现外电路电流输出(图5 ).获得离子浓差渗透能的关键是引入离子选择性膜,近年来研究较多的纳米通道浓差电池为植物发供电体系的设计提供了重要借鉴.浓差发电原理为在具有离子选择性的纳米通道两端建立离子浓度梯度,离子将从高浓度的一端向低浓度的一端扩散,由于孔道的离子选择性,阴阳离子通过孔道的流量不同,形成离子定向迁移的内电流,再通过阴极和阳极的电化学反应,将离子导体转变为电子导体(内电流转变为外电流),对负载进行供电[25 ] .因此,通过总结利用纳米通道实现离子浓度梯度发电的相关报道,本文期望可以将其原理和应用展望到植物体内,推动植物体离子浓度差发供电技术的发展. ...
1
... 植物在维持生命活动或应对环境变化时会自动调节各部位的离子浓度,从而为生命活动提供一个相对稳定的电解质环境以确保酶的活性[24 ] .植物体离子浓度差发供电技术就是利用植物不同部位的离子浓度不同,由离子浓度差异产生电势差实现外电路电流输出(图5 ).获得离子浓差渗透能的关键是引入离子选择性膜,近年来研究较多的纳米通道浓差电池为植物发供电体系的设计提供了重要借鉴.浓差发电原理为在具有离子选择性的纳米通道两端建立离子浓度梯度,离子将从高浓度的一端向低浓度的一端扩散,由于孔道的离子选择性,阴阳离子通过孔道的流量不同,形成离子定向迁移的内电流,再通过阴极和阳极的电化学反应,将离子导体转变为电子导体(内电流转变为外电流),对负载进行供电[25 ] .因此,通过总结利用纳米通道实现离子浓度梯度发电的相关报道,本文期望可以将其原理和应用展望到植物体内,推动植物体离子浓度差发供电技术的发展. ...
1
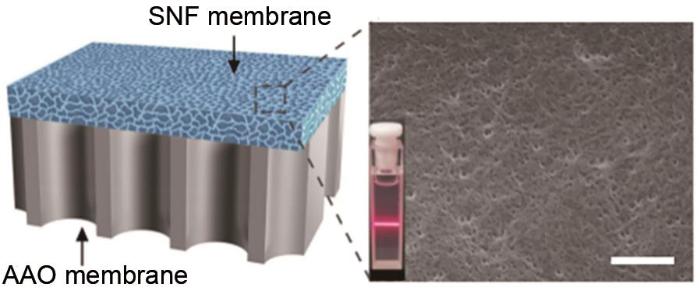
... 王亚等[26 ] 利用纳米多孔阳极氧化铝膜(AAO)实现了浓度梯度差发电,并研究了渗透电流、渗透电压和输出功率与浓度梯度差的关系.实验结果表明,渗透电压随着浓度梯度差的增大而增大,渗透电流随浓度梯度差的增大而逐渐减小,输出功率随着浓度梯度差的增加先增大后减小.该团队认为这是溶液浓度梯度差和纳米孔表面双电荷层厚度竞争的结果,并指出增加盐溶液的浓度虽然增加了浓度梯度差,提高了带电离子的扩散速率,但同时也会对纳米孔的离子选择透过性产生影响.此外,在最佳浓度梯度差下,用红墨水纳米颗粒对纳米多孔阳极氧化铝膜进行修饰,发现输出功率提高了近18倍,纳米孔阵列的离子选择透过性也有所增强.表明在相同的浓度梯度下,纳米孔阵列的离子选择透过性越强,输出功率越高.Xin等[27 ] 利用蚕丝纳米纤维膜(SNF)和阳极氧化铝膜(AAO)制备了一种具有不对称结构和电荷极性的高性能纳米流体复合膜,如图6 所示.SNF含有纳米通道,主导离子传输;AAO作为支撑基底,提供可调通道和两性基团.SNF表面有大量的羟基和氨基,AAO表面含有羟基,各基团之间形成氢键,增强了界面结合,赋予了复合膜优异的机械强度和稳定性.该团队探究了SNF的厚度和AAO通道的尺寸对能量转换性能的影响.实验结果表明,在不同的酸碱度条件下,SNF的最佳厚度为10~15 μm,AAO通道的最佳尺寸为80~100 nm.随着酸碱度从3.00增加到11.00,所有厚度和尺寸下的功率密度都有所增加,表明复合膜在碱性环境中表现出更好的性能.在50倍浓度梯度、负载电阻为23 kΩ时,复合膜的输出功率密度可达2.86 W/m2 . ...
1
... 王亚等[26 ] 利用纳米多孔阳极氧化铝膜(AAO)实现了浓度梯度差发电,并研究了渗透电流、渗透电压和输出功率与浓度梯度差的关系.实验结果表明,渗透电压随着浓度梯度差的增大而增大,渗透电流随浓度梯度差的增大而逐渐减小,输出功率随着浓度梯度差的增加先增大后减小.该团队认为这是溶液浓度梯度差和纳米孔表面双电荷层厚度竞争的结果,并指出增加盐溶液的浓度虽然增加了浓度梯度差,提高了带电离子的扩散速率,但同时也会对纳米孔的离子选择透过性产生影响.此外,在最佳浓度梯度差下,用红墨水纳米颗粒对纳米多孔阳极氧化铝膜进行修饰,发现输出功率提高了近18倍,纳米孔阵列的离子选择透过性也有所增强.表明在相同的浓度梯度下,纳米孔阵列的离子选择透过性越强,输出功率越高.Xin等[27 ] 利用蚕丝纳米纤维膜(SNF)和阳极氧化铝膜(AAO)制备了一种具有不对称结构和电荷极性的高性能纳米流体复合膜,如图6 所示.SNF含有纳米通道,主导离子传输;AAO作为支撑基底,提供可调通道和两性基团.SNF表面有大量的羟基和氨基,AAO表面含有羟基,各基团之间形成氢键,增强了界面结合,赋予了复合膜优异的机械强度和稳定性.该团队探究了SNF的厚度和AAO通道的尺寸对能量转换性能的影响.实验结果表明,在不同的酸碱度条件下,SNF的最佳厚度为10~15 μm,AAO通道的最佳尺寸为80~100 nm.随着酸碱度从3.00增加到11.00,所有厚度和尺寸下的功率密度都有所增加,表明复合膜在碱性环境中表现出更好的性能.在50倍浓度梯度、负载电阻为23 kΩ时,复合膜的输出功率密度可达2.86 W/m2 . ...
2
... 王亚等[26 ] 利用纳米多孔阳极氧化铝膜(AAO)实现了浓度梯度差发电,并研究了渗透电流、渗透电压和输出功率与浓度梯度差的关系.实验结果表明,渗透电压随着浓度梯度差的增大而增大,渗透电流随浓度梯度差的增大而逐渐减小,输出功率随着浓度梯度差的增加先增大后减小.该团队认为这是溶液浓度梯度差和纳米孔表面双电荷层厚度竞争的结果,并指出增加盐溶液的浓度虽然增加了浓度梯度差,提高了带电离子的扩散速率,但同时也会对纳米孔的离子选择透过性产生影响.此外,在最佳浓度梯度差下,用红墨水纳米颗粒对纳米多孔阳极氧化铝膜进行修饰,发现输出功率提高了近18倍,纳米孔阵列的离子选择透过性也有所增强.表明在相同的浓度梯度下,纳米孔阵列的离子选择透过性越强,输出功率越高.Xin等[27 ] 利用蚕丝纳米纤维膜(SNF)和阳极氧化铝膜(AAO)制备了一种具有不对称结构和电荷极性的高性能纳米流体复合膜,如图6 所示.SNF含有纳米通道,主导离子传输;AAO作为支撑基底,提供可调通道和两性基团.SNF表面有大量的羟基和氨基,AAO表面含有羟基,各基团之间形成氢键,增强了界面结合,赋予了复合膜优异的机械强度和稳定性.该团队探究了SNF的厚度和AAO通道的尺寸对能量转换性能的影响.实验结果表明,在不同的酸碱度条件下,SNF的最佳厚度为10~15 μm,AAO通道的最佳尺寸为80~100 nm.随着酸碱度从3.00增加到11.00,所有厚度和尺寸下的功率密度都有所增加,表明复合膜在碱性环境中表现出更好的性能.在50倍浓度梯度、负载电阻为23 kΩ时,复合膜的输出功率密度可达2.86 W/m2 . ...
... Summary of power generation performance parameters of concentration differential batteries based on ion-selective nanochannel
Table 1 二维纳米流体通道 膜厚度/μm 浓度梯度(倍数) 最大功率密度/(W/m2 ) 最大能量转换效率 参考文献 超薄氮化碳膜 0.25 1000 0.21 — [34 ] 二氧化硅膜 140 1000 7.70 31.0% [30 ] 异号电荷GO膜对 10 50 0.77 36.6% [35 ] 阳离子插层改性GO膜 — 1000 38.00 — [31 ] SPEEK膜 27 50 5.80 — [32 ] SNF/AAO复合膜 65 50 2.86 27.3% [27 ] SPEEK/AAO复合膜 6.5 50 4.80 — [36 ] Mxene/ANF复合膜 4.5 50 3.70 35.0% [28 ] 蒙脱土膜 70 1000 0.15 — [33 ] 离子二极管膜 — 50 3.46 37.3% [37 ]
综上所述,由离子选择性的纳米通道设计的浓差电池能够获得比较理想的输出功率密度,所以建立离子浓度梯度实现对外发供电具有理论可行性.该体系的设计需注意以下事项:植物叶片和组织内部的导管中存在含有离子浓度梯度的组织液,能够用于电流输出;电极需要嵌入植物本体,应采用惰性电极作为非牺牲电极,如铂电极、Ag/AgCl电极等,使电极不与组织液反应,获取持续的电流输出;能量输出强度依赖于离子浓差大小,由于发电过程的一部分在植物体内进行,所以能量输出还应受到植物的呼吸作用、蒸腾作用、光合作用以及外界环境变化的影响;植物的不同部位离子浓度梯度不同,选取不同部位进行发供电产生的能量输出应该不同.相较于牺牲电极的铜锌原电池,离子浓差电池的电极为惰性电极,不直接参与反应、不产生有毒物质,对植物生理过程干扰小,因此电池应具有更长的使用寿命,只要植物能正常维持生命活动就可以持续发供电.同时,植物体离子浓度差发供电的提出摆脱了水域的限制,不需要大量的具有浓度差异的液体作为供能条件,只要植物存在生命体征,就可以持续发供电,与传统浓差电池相比更具持久性. ...
2
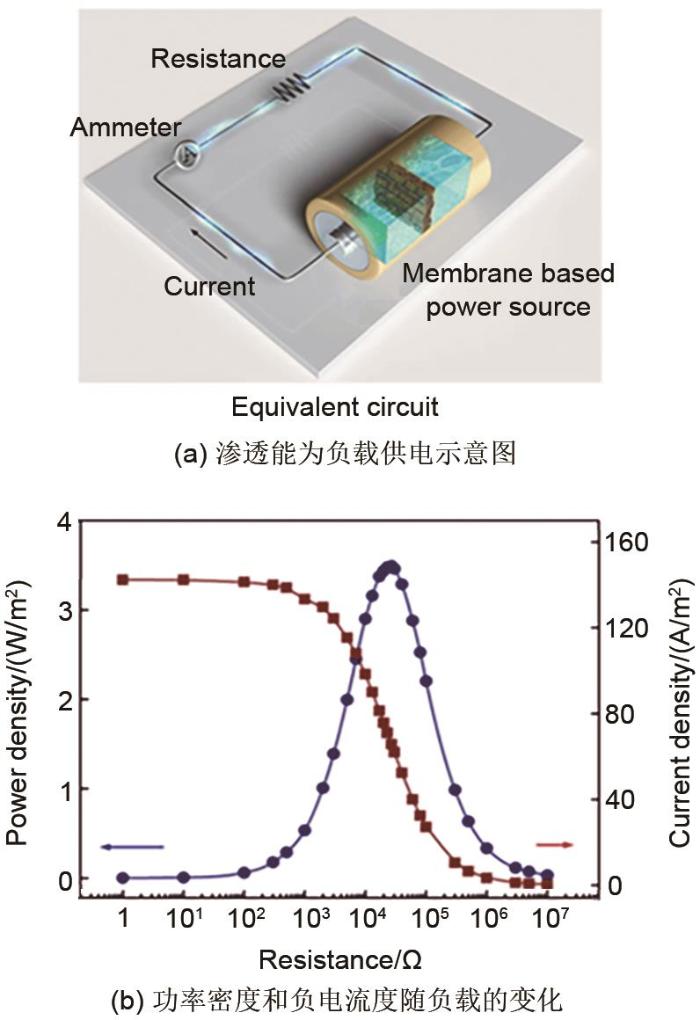
... 近期,Zhang等[28 ] 利用过渡金属碳化物(Mxene)和芳纶纳米纤维(ANF)制备了一种具有高机械强度和稳定性的Mxene/ANF复合膜,复合膜表现出良好的阳离子选择性和渗透能量转化性能,如图7 (a)所示.图7 (b)为ANF质量分数为11%的复合膜在人工海水(0.5 mol/L NaCl)和河水(0.01 mol/L NaCl)系统下的发电性能,电极为Ag/AgCl电极.随着电阻的增加,外部电路上的电流密度相应降低,输出功率密度在电阻为27 kΩ时达到最大值3.7 W/m2 .此外,复合膜在天然海水(0.6 mol/L NaCl)和河水(0.004 mol/L NaCl)系统下的最大功率密度可达约4.1 W/m2 ,更接近商业化基准(5 W/m2 ).在水溶液中,无机纳米通道的表面可以通过电离、离子吸附等途径获得电荷.这些表面电荷将吸引反离子,排斥同离子.存在浓度梯度时,反离子比同离子更容易通过纳米通道传输,导致离子的净电荷迁移[29 ] .因此可以使用无机离子选择性纳米通道将吉布斯自由能转化为电能.Kim等[30 ] 在硅片上制作了高度分别为4 nm、26 nm和80 nm的二氧化硅纳米通道(长140 μm,宽25 μm,由这些纳米通道制成的平行纳米通道阵列的孔隙率为0.25),通过实验研究无机纳米通道在不同浓度KCl溶液之间的发电情况,所用电极为Ag/AgCl电极.实验结果表明,单位体积二氧化硅纳米通道的发电量随着浓度梯度的增加而增加,同时随着通道高度的降低而增加,测得的最高功率密度为7.7 W/m2 .此外,二氧化硅纳米通道的能量转换效率随着浓度梯度和通道高度的降低而增加,获得的最佳能量转换效率为31%,与一些商用能量转换膜相当.无机离子选择性纳米通道中的浓度梯度发电可用于各种实际应用,包括微型电池和微型发电机等. ...
... Summary of power generation performance parameters of concentration differential batteries based on ion-selective nanochannel
Table 1 二维纳米流体通道 膜厚度/μm 浓度梯度(倍数) 最大功率密度/(W/m2 ) 最大能量转换效率 参考文献 超薄氮化碳膜 0.25 1000 0.21 — [34 ] 二氧化硅膜 140 1000 7.70 31.0% [30 ] 异号电荷GO膜对 10 50 0.77 36.6% [35 ] 阳离子插层改性GO膜 — 1000 38.00 — [31 ] SPEEK膜 27 50 5.80 — [32 ] SNF/AAO复合膜 65 50 2.86 27.3% [27 ] SPEEK/AAO复合膜 6.5 50 4.80 — [36 ] Mxene/ANF复合膜 4.5 50 3.70 35.0% [28 ] 蒙脱土膜 70 1000 0.15 — [33 ] 离子二极管膜 — 50 3.46 37.3% [37 ]
综上所述,由离子选择性的纳米通道设计的浓差电池能够获得比较理想的输出功率密度,所以建立离子浓度梯度实现对外发供电具有理论可行性.该体系的设计需注意以下事项:植物叶片和组织内部的导管中存在含有离子浓度梯度的组织液,能够用于电流输出;电极需要嵌入植物本体,应采用惰性电极作为非牺牲电极,如铂电极、Ag/AgCl电极等,使电极不与组织液反应,获取持续的电流输出;能量输出强度依赖于离子浓差大小,由于发电过程的一部分在植物体内进行,所以能量输出还应受到植物的呼吸作用、蒸腾作用、光合作用以及外界环境变化的影响;植物的不同部位离子浓度梯度不同,选取不同部位进行发供电产生的能量输出应该不同.相较于牺牲电极的铜锌原电池,离子浓差电池的电极为惰性电极,不直接参与反应、不产生有毒物质,对植物生理过程干扰小,因此电池应具有更长的使用寿命,只要植物能正常维持生命活动就可以持续发供电.同时,植物体离子浓度差发供电的提出摆脱了水域的限制,不需要大量的具有浓度差异的液体作为供能条件,只要植物存在生命体征,就可以持续发供电,与传统浓差电池相比更具持久性. ...
1
... 近期,Zhang等[28 ] 利用过渡金属碳化物(Mxene)和芳纶纳米纤维(ANF)制备了一种具有高机械强度和稳定性的Mxene/ANF复合膜,复合膜表现出良好的阳离子选择性和渗透能量转化性能,如图7 (a)所示.图7 (b)为ANF质量分数为11%的复合膜在人工海水(0.5 mol/L NaCl)和河水(0.01 mol/L NaCl)系统下的发电性能,电极为Ag/AgCl电极.随着电阻的增加,外部电路上的电流密度相应降低,输出功率密度在电阻为27 kΩ时达到最大值3.7 W/m2 .此外,复合膜在天然海水(0.6 mol/L NaCl)和河水(0.004 mol/L NaCl)系统下的最大功率密度可达约4.1 W/m2 ,更接近商业化基准(5 W/m2 ).在水溶液中,无机纳米通道的表面可以通过电离、离子吸附等途径获得电荷.这些表面电荷将吸引反离子,排斥同离子.存在浓度梯度时,反离子比同离子更容易通过纳米通道传输,导致离子的净电荷迁移[29 ] .因此可以使用无机离子选择性纳米通道将吉布斯自由能转化为电能.Kim等[30 ] 在硅片上制作了高度分别为4 nm、26 nm和80 nm的二氧化硅纳米通道(长140 μm,宽25 μm,由这些纳米通道制成的平行纳米通道阵列的孔隙率为0.25),通过实验研究无机纳米通道在不同浓度KCl溶液之间的发电情况,所用电极为Ag/AgCl电极.实验结果表明,单位体积二氧化硅纳米通道的发电量随着浓度梯度的增加而增加,同时随着通道高度的降低而增加,测得的最高功率密度为7.7 W/m2 .此外,二氧化硅纳米通道的能量转换效率随着浓度梯度和通道高度的降低而增加,获得的最佳能量转换效率为31%,与一些商用能量转换膜相当.无机离子选择性纳米通道中的浓度梯度发电可用于各种实际应用,包括微型电池和微型发电机等. ...
2
... 近期,Zhang等[28 ] 利用过渡金属碳化物(Mxene)和芳纶纳米纤维(ANF)制备了一种具有高机械强度和稳定性的Mxene/ANF复合膜,复合膜表现出良好的阳离子选择性和渗透能量转化性能,如图7 (a)所示.图7 (b)为ANF质量分数为11%的复合膜在人工海水(0.5 mol/L NaCl)和河水(0.01 mol/L NaCl)系统下的发电性能,电极为Ag/AgCl电极.随着电阻的增加,外部电路上的电流密度相应降低,输出功率密度在电阻为27 kΩ时达到最大值3.7 W/m2 .此外,复合膜在天然海水(0.6 mol/L NaCl)和河水(0.004 mol/L NaCl)系统下的最大功率密度可达约4.1 W/m2 ,更接近商业化基准(5 W/m2 ).在水溶液中,无机纳米通道的表面可以通过电离、离子吸附等途径获得电荷.这些表面电荷将吸引反离子,排斥同离子.存在浓度梯度时,反离子比同离子更容易通过纳米通道传输,导致离子的净电荷迁移[29 ] .因此可以使用无机离子选择性纳米通道将吉布斯自由能转化为电能.Kim等[30 ] 在硅片上制作了高度分别为4 nm、26 nm和80 nm的二氧化硅纳米通道(长140 μm,宽25 μm,由这些纳米通道制成的平行纳米通道阵列的孔隙率为0.25),通过实验研究无机纳米通道在不同浓度KCl溶液之间的发电情况,所用电极为Ag/AgCl电极.实验结果表明,单位体积二氧化硅纳米通道的发电量随着浓度梯度的增加而增加,同时随着通道高度的降低而增加,测得的最高功率密度为7.7 W/m2 .此外,二氧化硅纳米通道的能量转换效率随着浓度梯度和通道高度的降低而增加,获得的最佳能量转换效率为31%,与一些商用能量转换膜相当.无机离子选择性纳米通道中的浓度梯度发电可用于各种实际应用,包括微型电池和微型发电机等. ...
... Summary of power generation performance parameters of concentration differential batteries based on ion-selective nanochannel
Table 1 二维纳米流体通道 膜厚度/μm 浓度梯度(倍数) 最大功率密度/(W/m2 ) 最大能量转换效率 参考文献 超薄氮化碳膜 0.25 1000 0.21 — [34 ] 二氧化硅膜 140 1000 7.70 31.0% [30 ] 异号电荷GO膜对 10 50 0.77 36.6% [35 ] 阳离子插层改性GO膜 — 1000 38.00 — [31 ] SPEEK膜 27 50 5.80 — [32 ] SNF/AAO复合膜 65 50 2.86 27.3% [27 ] SPEEK/AAO复合膜 6.5 50 4.80 — [36 ] Mxene/ANF复合膜 4.5 50 3.70 35.0% [28 ] 蒙脱土膜 70 1000 0.15 — [33 ] 离子二极管膜 — 50 3.46 37.3% [37 ]
综上所述,由离子选择性的纳米通道设计的浓差电池能够获得比较理想的输出功率密度,所以建立离子浓度梯度实现对外发供电具有理论可行性.该体系的设计需注意以下事项:植物叶片和组织内部的导管中存在含有离子浓度梯度的组织液,能够用于电流输出;电极需要嵌入植物本体,应采用惰性电极作为非牺牲电极,如铂电极、Ag/AgCl电极等,使电极不与组织液反应,获取持续的电流输出;能量输出强度依赖于离子浓差大小,由于发电过程的一部分在植物体内进行,所以能量输出还应受到植物的呼吸作用、蒸腾作用、光合作用以及外界环境变化的影响;植物的不同部位离子浓度梯度不同,选取不同部位进行发供电产生的能量输出应该不同.相较于牺牲电极的铜锌原电池,离子浓差电池的电极为惰性电极,不直接参与反应、不产生有毒物质,对植物生理过程干扰小,因此电池应具有更长的使用寿命,只要植物能正常维持生命活动就可以持续发供电.同时,植物体离子浓度差发供电的提出摆脱了水域的限制,不需要大量的具有浓度差异的液体作为供能条件,只要植物存在生命体征,就可以持续发供电,与传统浓差电池相比更具持久性. ...
2
... 氧化石墨烯纤维膜由于其孔径大小和层间距离可以调控,在能量转换等方面表现优异,可以用于盐度梯度发电.然而,氧化石墨烯纤维膜处在水溶液中时,其层间间距会发生膨胀,削弱了限制离子传输的能力,不能获得足够的功率密度(低于1 W/m2 ).为此,Ghanbari等[31 ] 制备了阳离子插层的改性氧化石墨烯纤维膜,根据阳离子的水合尺寸控制中间层的间距,这种对层间间距的控制导致具有较大水合体积的阳离子被排除在外,而较小的阳离子仍然可以通过,从而增强对离子传输的稳定限制.在1000倍的氯化钾浓度梯度下,负载电阻为360 kΩ时,产生的渗透功率密度达到了38 W/m2 ,远远超过了传统的氧化石墨烯纤维膜,具有优异的可扩展性和稳定性.离子选择性纳米通道薄膜很好地推动了盐度梯度发电技术的发展,但是大多数薄膜制备复杂、机械强度不高且功率密度输出不能满足商业化标准(5 W/m2 ),限制了盐度梯度发电的大规模应用.为此,Zhao等[32 ] 通过非溶剂诱导相分离的方法制备了含有大量纳米通道的磺化聚醚醚酮(SPEEK)膜,用于将盐度梯度能量转化为电能.该膜由海绵状多孔支撑层和富含平行连续纳米通道的致密表层构成,纳米通道通过促进阳离子的输运和排斥阴离子来控制离子选择性传输.通过混合人工海水和河水,基于该膜的供电装置的输出功率密度达到5.8 W/m2 ,满足商业化基准.功率密度还可以通过调节膜的厚度和磺化度来进一步提高.此外,SPEEK膜表现出优异的力学性能和高耐久性,使其有可能建设实际的盐度梯度发电系统,用于商业化. ...
... Summary of power generation performance parameters of concentration differential batteries based on ion-selective nanochannel
Table 1 二维纳米流体通道 膜厚度/μm 浓度梯度(倍数) 最大功率密度/(W/m2 ) 最大能量转换效率 参考文献 超薄氮化碳膜 0.25 1000 0.21 — [34 ] 二氧化硅膜 140 1000 7.70 31.0% [30 ] 异号电荷GO膜对 10 50 0.77 36.6% [35 ] 阳离子插层改性GO膜 — 1000 38.00 — [31 ] SPEEK膜 27 50 5.80 — [32 ] SNF/AAO复合膜 65 50 2.86 27.3% [27 ] SPEEK/AAO复合膜 6.5 50 4.80 — [36 ] Mxene/ANF复合膜 4.5 50 3.70 35.0% [28 ] 蒙脱土膜 70 1000 0.15 — [33 ] 离子二极管膜 — 50 3.46 37.3% [37 ]
综上所述,由离子选择性的纳米通道设计的浓差电池能够获得比较理想的输出功率密度,所以建立离子浓度梯度实现对外发供电具有理论可行性.该体系的设计需注意以下事项:植物叶片和组织内部的导管中存在含有离子浓度梯度的组织液,能够用于电流输出;电极需要嵌入植物本体,应采用惰性电极作为非牺牲电极,如铂电极、Ag/AgCl电极等,使电极不与组织液反应,获取持续的电流输出;能量输出强度依赖于离子浓差大小,由于发电过程的一部分在植物体内进行,所以能量输出还应受到植物的呼吸作用、蒸腾作用、光合作用以及外界环境变化的影响;植物的不同部位离子浓度梯度不同,选取不同部位进行发供电产生的能量输出应该不同.相较于牺牲电极的铜锌原电池,离子浓差电池的电极为惰性电极,不直接参与反应、不产生有毒物质,对植物生理过程干扰小,因此电池应具有更长的使用寿命,只要植物能正常维持生命活动就可以持续发供电.同时,植物体离子浓度差发供电的提出摆脱了水域的限制,不需要大量的具有浓度差异的液体作为供能条件,只要植物存在生命体征,就可以持续发供电,与传统浓差电池相比更具持久性. ...
2
... 氧化石墨烯纤维膜由于其孔径大小和层间距离可以调控,在能量转换等方面表现优异,可以用于盐度梯度发电.然而,氧化石墨烯纤维膜处在水溶液中时,其层间间距会发生膨胀,削弱了限制离子传输的能力,不能获得足够的功率密度(低于1 W/m2 ).为此,Ghanbari等[31 ] 制备了阳离子插层的改性氧化石墨烯纤维膜,根据阳离子的水合尺寸控制中间层的间距,这种对层间间距的控制导致具有较大水合体积的阳离子被排除在外,而较小的阳离子仍然可以通过,从而增强对离子传输的稳定限制.在1000倍的氯化钾浓度梯度下,负载电阻为360 kΩ时,产生的渗透功率密度达到了38 W/m2 ,远远超过了传统的氧化石墨烯纤维膜,具有优异的可扩展性和稳定性.离子选择性纳米通道薄膜很好地推动了盐度梯度发电技术的发展,但是大多数薄膜制备复杂、机械强度不高且功率密度输出不能满足商业化标准(5 W/m2 ),限制了盐度梯度发电的大规模应用.为此,Zhao等[32 ] 通过非溶剂诱导相分离的方法制备了含有大量纳米通道的磺化聚醚醚酮(SPEEK)膜,用于将盐度梯度能量转化为电能.该膜由海绵状多孔支撑层和富含平行连续纳米通道的致密表层构成,纳米通道通过促进阳离子的输运和排斥阴离子来控制离子选择性传输.通过混合人工海水和河水,基于该膜的供电装置的输出功率密度达到5.8 W/m2 ,满足商业化基准.功率密度还可以通过调节膜的厚度和磺化度来进一步提高.此外,SPEEK膜表现出优异的力学性能和高耐久性,使其有可能建设实际的盐度梯度发电系统,用于商业化. ...
... Summary of power generation performance parameters of concentration differential batteries based on ion-selective nanochannel
Table 1 二维纳米流体通道 膜厚度/μm 浓度梯度(倍数) 最大功率密度/(W/m2 ) 最大能量转换效率 参考文献 超薄氮化碳膜 0.25 1000 0.21 — [34 ] 二氧化硅膜 140 1000 7.70 31.0% [30 ] 异号电荷GO膜对 10 50 0.77 36.6% [35 ] 阳离子插层改性GO膜 — 1000 38.00 — [31 ] SPEEK膜 27 50 5.80 — [32 ] SNF/AAO复合膜 65 50 2.86 27.3% [27 ] SPEEK/AAO复合膜 6.5 50 4.80 — [36 ] Mxene/ANF复合膜 4.5 50 3.70 35.0% [28 ] 蒙脱土膜 70 1000 0.15 — [33 ] 离子二极管膜 — 50 3.46 37.3% [37 ]
综上所述,由离子选择性的纳米通道设计的浓差电池能够获得比较理想的输出功率密度,所以建立离子浓度梯度实现对外发供电具有理论可行性.该体系的设计需注意以下事项:植物叶片和组织内部的导管中存在含有离子浓度梯度的组织液,能够用于电流输出;电极需要嵌入植物本体,应采用惰性电极作为非牺牲电极,如铂电极、Ag/AgCl电极等,使电极不与组织液反应,获取持续的电流输出;能量输出强度依赖于离子浓差大小,由于发电过程的一部分在植物体内进行,所以能量输出还应受到植物的呼吸作用、蒸腾作用、光合作用以及外界环境变化的影响;植物的不同部位离子浓度梯度不同,选取不同部位进行发供电产生的能量输出应该不同.相较于牺牲电极的铜锌原电池,离子浓差电池的电极为惰性电极,不直接参与反应、不产生有毒物质,对植物生理过程干扰小,因此电池应具有更长的使用寿命,只要植物能正常维持生命活动就可以持续发供电.同时,植物体离子浓度差发供电的提出摆脱了水域的限制,不需要大量的具有浓度差异的液体作为供能条件,只要植物存在生命体征,就可以持续发供电,与传统浓差电池相比更具持久性. ...
2
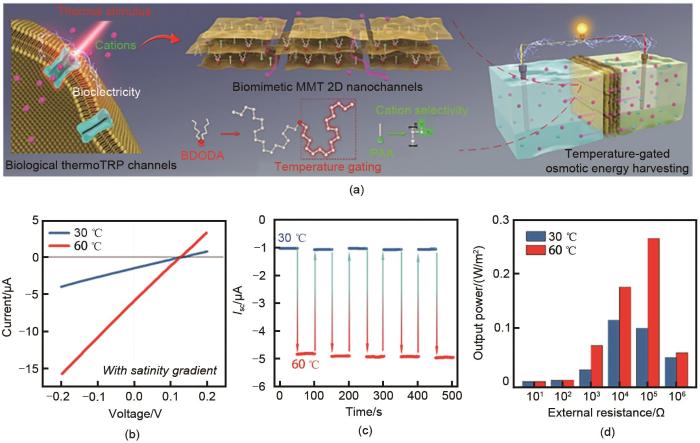
... 此外,纳米通道浓差电池的能量输出可以通过外场进行调控.本课题组设计了由功能化蒙脱土(MMT)层堆集而成的温度控制二维阳离子纳米通道[33 ] ,用于实现浓差电池的可控能量输出,如图8 (a)所示.纳米通道整合了两个功能部分,一种是聚丙烯酸(PAA)分子,其在纳米通道表面产生高密度的负电荷,以获得优异的阳离子选择性.另一种是双十八烷基二甲基溴化铵(DODAB,相变温度约为45 ℃),其在不同温度之间的构型转变有助于阳离子门控行为.图8 (b)为浓差电池在1000倍盐度梯度下的伏安特性曲线,在60 ℃时,浓差电池的电流输出有所提高,这表明了温度可控的能量输出.在图8 (c)所示的温度周期转换中,短路电流的变化表现出良好的可逆性和稳定性.图8 (d)为不同温度和外部电阻下的输出功率密度情况.随着温度从30 ℃增加到60 ℃,外部电阻负载的输出功率密度增加,峰值为0.34 W/m2 .外部电阻为105 Ω时,输出功率密度相差两倍以上.基于阳离子选择性和温度控制离子传输,纳米通道浓差电池可以通过温度的交替变化,可逆稳定地调节能量输出,为能源的可控利用提供新的途径. ...
... Summary of power generation performance parameters of concentration differential batteries based on ion-selective nanochannel
Table 1 二维纳米流体通道 膜厚度/μm 浓度梯度(倍数) 最大功率密度/(W/m2 ) 最大能量转换效率 参考文献 超薄氮化碳膜 0.25 1000 0.21 — [34 ] 二氧化硅膜 140 1000 7.70 31.0% [30 ] 异号电荷GO膜对 10 50 0.77 36.6% [35 ] 阳离子插层改性GO膜 — 1000 38.00 — [31 ] SPEEK膜 27 50 5.80 — [32 ] SNF/AAO复合膜 65 50 2.86 27.3% [27 ] SPEEK/AAO复合膜 6.5 50 4.80 — [36 ] Mxene/ANF复合膜 4.5 50 3.70 35.0% [28 ] 蒙脱土膜 70 1000 0.15 — [33 ] 离子二极管膜 — 50 3.46 37.3% [37 ]
综上所述,由离子选择性的纳米通道设计的浓差电池能够获得比较理想的输出功率密度,所以建立离子浓度梯度实现对外发供电具有理论可行性.该体系的设计需注意以下事项:植物叶片和组织内部的导管中存在含有离子浓度梯度的组织液,能够用于电流输出;电极需要嵌入植物本体,应采用惰性电极作为非牺牲电极,如铂电极、Ag/AgCl电极等,使电极不与组织液反应,获取持续的电流输出;能量输出强度依赖于离子浓差大小,由于发电过程的一部分在植物体内进行,所以能量输出还应受到植物的呼吸作用、蒸腾作用、光合作用以及外界环境变化的影响;植物的不同部位离子浓度梯度不同,选取不同部位进行发供电产生的能量输出应该不同.相较于牺牲电极的铜锌原电池,离子浓差电池的电极为惰性电极,不直接参与反应、不产生有毒物质,对植物生理过程干扰小,因此电池应具有更长的使用寿命,只要植物能正常维持生命活动就可以持续发供电.同时,植物体离子浓度差发供电的提出摆脱了水域的限制,不需要大量的具有浓度差异的液体作为供能条件,只要植物存在生命体征,就可以持续发供电,与传统浓差电池相比更具持久性. ...
1
... Summary of power generation performance parameters of concentration differential batteries based on ion-selective nanochannel
Table 1 二维纳米流体通道 膜厚度/μm 浓度梯度(倍数) 最大功率密度/(W/m2 ) 最大能量转换效率 参考文献 超薄氮化碳膜 0.25 1000 0.21 — [34 ] 二氧化硅膜 140 1000 7.70 31.0% [30 ] 异号电荷GO膜对 10 50 0.77 36.6% [35 ] 阳离子插层改性GO膜 — 1000 38.00 — [31 ] SPEEK膜 27 50 5.80 — [32 ] SNF/AAO复合膜 65 50 2.86 27.3% [27 ] SPEEK/AAO复合膜 6.5 50 4.80 — [36 ] Mxene/ANF复合膜 4.5 50 3.70 35.0% [28 ] 蒙脱土膜 70 1000 0.15 — [33 ] 离子二极管膜 — 50 3.46 37.3% [37 ]
综上所述,由离子选择性的纳米通道设计的浓差电池能够获得比较理想的输出功率密度,所以建立离子浓度梯度实现对外发供电具有理论可行性.该体系的设计需注意以下事项:植物叶片和组织内部的导管中存在含有离子浓度梯度的组织液,能够用于电流输出;电极需要嵌入植物本体,应采用惰性电极作为非牺牲电极,如铂电极、Ag/AgCl电极等,使电极不与组织液反应,获取持续的电流输出;能量输出强度依赖于离子浓差大小,由于发电过程的一部分在植物体内进行,所以能量输出还应受到植物的呼吸作用、蒸腾作用、光合作用以及外界环境变化的影响;植物的不同部位离子浓度梯度不同,选取不同部位进行发供电产生的能量输出应该不同.相较于牺牲电极的铜锌原电池,离子浓差电池的电极为惰性电极,不直接参与反应、不产生有毒物质,对植物生理过程干扰小,因此电池应具有更长的使用寿命,只要植物能正常维持生命活动就可以持续发供电.同时,植物体离子浓度差发供电的提出摆脱了水域的限制,不需要大量的具有浓度差异的液体作为供能条件,只要植物存在生命体征,就可以持续发供电,与传统浓差电池相比更具持久性. ...
1
... Summary of power generation performance parameters of concentration differential batteries based on ion-selective nanochannel
Table 1 二维纳米流体通道 膜厚度/μm 浓度梯度(倍数) 最大功率密度/(W/m2 ) 最大能量转换效率 参考文献 超薄氮化碳膜 0.25 1000 0.21 — [34 ] 二氧化硅膜 140 1000 7.70 31.0% [30 ] 异号电荷GO膜对 10 50 0.77 36.6% [35 ] 阳离子插层改性GO膜 — 1000 38.00 — [31 ] SPEEK膜 27 50 5.80 — [32 ] SNF/AAO复合膜 65 50 2.86 27.3% [27 ] SPEEK/AAO复合膜 6.5 50 4.80 — [36 ] Mxene/ANF复合膜 4.5 50 3.70 35.0% [28 ] 蒙脱土膜 70 1000 0.15 — [33 ] 离子二极管膜 — 50 3.46 37.3% [37 ]
综上所述,由离子选择性的纳米通道设计的浓差电池能够获得比较理想的输出功率密度,所以建立离子浓度梯度实现对外发供电具有理论可行性.该体系的设计需注意以下事项:植物叶片和组织内部的导管中存在含有离子浓度梯度的组织液,能够用于电流输出;电极需要嵌入植物本体,应采用惰性电极作为非牺牲电极,如铂电极、Ag/AgCl电极等,使电极不与组织液反应,获取持续的电流输出;能量输出强度依赖于离子浓差大小,由于发电过程的一部分在植物体内进行,所以能量输出还应受到植物的呼吸作用、蒸腾作用、光合作用以及外界环境变化的影响;植物的不同部位离子浓度梯度不同,选取不同部位进行发供电产生的能量输出应该不同.相较于牺牲电极的铜锌原电池,离子浓差电池的电极为惰性电极,不直接参与反应、不产生有毒物质,对植物生理过程干扰小,因此电池应具有更长的使用寿命,只要植物能正常维持生命活动就可以持续发供电.同时,植物体离子浓度差发供电的提出摆脱了水域的限制,不需要大量的具有浓度差异的液体作为供能条件,只要植物存在生命体征,就可以持续发供电,与传统浓差电池相比更具持久性. ...
1
... Summary of power generation performance parameters of concentration differential batteries based on ion-selective nanochannel
Table 1 二维纳米流体通道 膜厚度/μm 浓度梯度(倍数) 最大功率密度/(W/m2 ) 最大能量转换效率 参考文献 超薄氮化碳膜 0.25 1000 0.21 — [34 ] 二氧化硅膜 140 1000 7.70 31.0% [30 ] 异号电荷GO膜对 10 50 0.77 36.6% [35 ] 阳离子插层改性GO膜 — 1000 38.00 — [31 ] SPEEK膜 27 50 5.80 — [32 ] SNF/AAO复合膜 65 50 2.86 27.3% [27 ] SPEEK/AAO复合膜 6.5 50 4.80 — [36 ] Mxene/ANF复合膜 4.5 50 3.70 35.0% [28 ] 蒙脱土膜 70 1000 0.15 — [33 ] 离子二极管膜 — 50 3.46 37.3% [37 ]
综上所述,由离子选择性的纳米通道设计的浓差电池能够获得比较理想的输出功率密度,所以建立离子浓度梯度实现对外发供电具有理论可行性.该体系的设计需注意以下事项:植物叶片和组织内部的导管中存在含有离子浓度梯度的组织液,能够用于电流输出;电极需要嵌入植物本体,应采用惰性电极作为非牺牲电极,如铂电极、Ag/AgCl电极等,使电极不与组织液反应,获取持续的电流输出;能量输出强度依赖于离子浓差大小,由于发电过程的一部分在植物体内进行,所以能量输出还应受到植物的呼吸作用、蒸腾作用、光合作用以及外界环境变化的影响;植物的不同部位离子浓度梯度不同,选取不同部位进行发供电产生的能量输出应该不同.相较于牺牲电极的铜锌原电池,离子浓差电池的电极为惰性电极,不直接参与反应、不产生有毒物质,对植物生理过程干扰小,因此电池应具有更长的使用寿命,只要植物能正常维持生命活动就可以持续发供电.同时,植物体离子浓度差发供电的提出摆脱了水域的限制,不需要大量的具有浓度差异的液体作为供能条件,只要植物存在生命体征,就可以持续发供电,与传统浓差电池相比更具持久性. ...
1
... Summary of power generation performance parameters of concentration differential batteries based on ion-selective nanochannel
Table 1 二维纳米流体通道 膜厚度/μm 浓度梯度(倍数) 最大功率密度/(W/m2 ) 最大能量转换效率 参考文献 超薄氮化碳膜 0.25 1000 0.21 — [34 ] 二氧化硅膜 140 1000 7.70 31.0% [30 ] 异号电荷GO膜对 10 50 0.77 36.6% [35 ] 阳离子插层改性GO膜 — 1000 38.00 — [31 ] SPEEK膜 27 50 5.80 — [32 ] SNF/AAO复合膜 65 50 2.86 27.3% [27 ] SPEEK/AAO复合膜 6.5 50 4.80 — [36 ] Mxene/ANF复合膜 4.5 50 3.70 35.0% [28 ] 蒙脱土膜 70 1000 0.15 — [33 ] 离子二极管膜 — 50 3.46 37.3% [37 ]
综上所述,由离子选择性的纳米通道设计的浓差电池能够获得比较理想的输出功率密度,所以建立离子浓度梯度实现对外发供电具有理论可行性.该体系的设计需注意以下事项:植物叶片和组织内部的导管中存在含有离子浓度梯度的组织液,能够用于电流输出;电极需要嵌入植物本体,应采用惰性电极作为非牺牲电极,如铂电极、Ag/AgCl电极等,使电极不与组织液反应,获取持续的电流输出;能量输出强度依赖于离子浓差大小,由于发电过程的一部分在植物体内进行,所以能量输出还应受到植物的呼吸作用、蒸腾作用、光合作用以及外界环境变化的影响;植物的不同部位离子浓度梯度不同,选取不同部位进行发供电产生的能量输出应该不同.相较于牺牲电极的铜锌原电池,离子浓差电池的电极为惰性电极,不直接参与反应、不产生有毒物质,对植物生理过程干扰小,因此电池应具有更长的使用寿命,只要植物能正常维持生命活动就可以持续发供电.同时,植物体离子浓度差发供电的提出摆脱了水域的限制,不需要大量的具有浓度差异的液体作为供能条件,只要植物存在生命体征,就可以持续发供电,与传统浓差电池相比更具持久性. ...
1
... 光合作用是指绿色植物吸收光能,把二氧化碳和水合成含能有机物,同时释放氧气的过程,是自然界中利用太阳能最普遍的方式,对实现自然界的能量转换、维持大气的碳氧平衡具有重要意义[38 ] .类光合作用发供电技术是通过电化学手段模拟或部分模拟光合作用等生物高级作用过程,通过电极将反应过程中的电子定向导出实现发供电的技术,该技术成本较低且环保性好,目前正在被广泛研究. ...
1
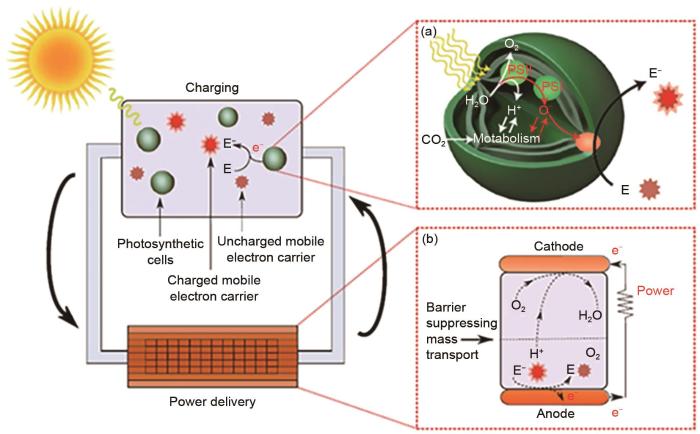
... 类光合作用发供电技术中最典型的是生物光伏技术(BPV)[39 ] ,其主要利用含氧光合生物来收集光能并提供电能输出.在光营养生物中,光能被转换成电荷分离的电子空穴对,受激发的电子通过细胞内的电子载体被转移,其中一部分最终通过细胞膜出口被释放到外部环境中,如图9 (a)所示.在生物光电池中,这些电子被引导到阳极,然后通过外部电路流向阴极,从而产生电流.同时,电池释放的质子从阳极室扩散到阴极室,在阴极室催化剂的作用下产生水,如图9 (b)所示,该过程产生电流而不释放任何化学副产物[40 ] . ...
1
... 类光合作用发供电技术中最典型的是生物光伏技术(BPV)[39 ] ,其主要利用含氧光合生物来收集光能并提供电能输出.在光营养生物中,光能被转换成电荷分离的电子空穴对,受激发的电子通过细胞内的电子载体被转移,其中一部分最终通过细胞膜出口被释放到外部环境中,如图9 (a)所示.在生物光电池中,这些电子被引导到阳极,然后通过外部电路流向阴极,从而产生电流.同时,电池释放的质子从阳极室扩散到阴极室,在阴极室催化剂的作用下产生水,如图9 (b)所示,该过程产生电流而不释放任何化学副产物[40 ] . ...
1
... 生物光伏技术提供的功率输出较低,一般比太阳能光伏电池低3个数量级以上[41 ] .为此,英国剑桥大学的Bombelli教授团队[42 ] 设计了一个实验平台,利用微型结构来减少电荷传输的距离,从而提供较大的功率输出.该团队开发了一种软光刻方法,用于制造不需要膜或介质的微流体生物光伏器件.通过将聚胞菌sp. PCC 6803细胞注入并使其沉淀在阳极上,使细胞与电极之间的物理接触达到无介质要求.在白光条件下,当叶绿素浓度为10-4 mol Chl/L时,功率密度超过100 mW/m2 ,这对于没有外部能量供应的生物光伏器件来说是一个很高的数值. ...
1
... 生物光伏技术提供的功率输出较低,一般比太阳能光伏电池低3个数量级以上[41 ] .为此,英国剑桥大学的Bombelli教授团队[42 ] 设计了一个实验平台,利用微型结构来减少电荷传输的距离,从而提供较大的功率输出.该团队开发了一种软光刻方法,用于制造不需要膜或介质的微流体生物光伏器件.通过将聚胞菌sp. PCC 6803细胞注入并使其沉淀在阳极上,使细胞与电极之间的物理接触达到无介质要求.在白光条件下,当叶绿素浓度为10-4 mol Chl/L时,功率密度超过100 mW/m2 ,这对于没有外部能量供应的生物光伏器件来说是一个很高的数值. ...
1
... 蓝藻是光合作用效率最高的生物之一,它们是原核生物,能量转换在类囊体膜中进行[43 ] .为了研究藻类用于制备生物光伏器件的能力,Ng等[44 ] 在氧化铟锡阳极和还原氧化石墨烯阳极上分别制备了长聚球藻的藻类生物膜,用于在光照和黑暗条件下研究藻类生物光伏器件.研究证明,与以氧化铟锡为阳极的生物光伏器件相比,以还原氧化石墨烯为阳极的生物光伏器件具有更高的功率输出,其功率密度最高可达0.538 mW/m2 ±0.014 mW/m2 ,说明了藻类生物光伏电池在光照下具有产生生物电的能力.Zhu等[45 ] 开发了一种具有定向电子流的以D-乳酸为中介的微生物组生物光伏技术,来提高光蓝藻的细胞外电子传输能力,从而提高BPV的效率和使用寿命,如图10 所示.该微生物组由能够将光能储存在D-乳酸的蓝藻和能够高效利用D-乳酸产电的希瓦菌组成,其中D-乳酸是两种微生物间的能量载体.蓝藻能够吸收光能并固定CO2 来合成能量载体D-乳酸,希瓦菌氧化D-乳酸进行产电,由此形成一条从光子到D-乳酸再到电能的定向电子流,完成从光能到化学能再到电能的能量转化过程.研究结果表明,该双菌生物光伏系统可以实现稳定、高效的功率输出,功率密度最高可达150 mW/m2 ,比普通单菌光伏系统高出10倍以上.该生物光伏技术可以使具有介质补充功能的装置稳定运行40 d以上,平均功率密度为135 mW/m2 ,在产电时长和功率输出方面均达到了较高水平.说明具有定向电子流的微生物组可以促进电子从光合细胞输出,并获得高效和持久的功率输出. ...
1
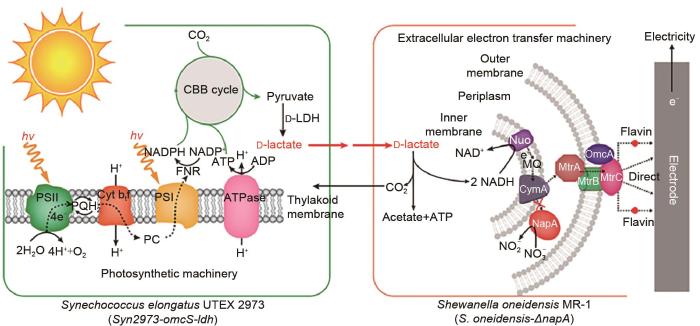
... 蓝藻是光合作用效率最高的生物之一,它们是原核生物,能量转换在类囊体膜中进行[43 ] .为了研究藻类用于制备生物光伏器件的能力,Ng等[44 ] 在氧化铟锡阳极和还原氧化石墨烯阳极上分别制备了长聚球藻的藻类生物膜,用于在光照和黑暗条件下研究藻类生物光伏器件.研究证明,与以氧化铟锡为阳极的生物光伏器件相比,以还原氧化石墨烯为阳极的生物光伏器件具有更高的功率输出,其功率密度最高可达0.538 mW/m2 ±0.014 mW/m2 ,说明了藻类生物光伏电池在光照下具有产生生物电的能力.Zhu等[45 ] 开发了一种具有定向电子流的以D-乳酸为中介的微生物组生物光伏技术,来提高光蓝藻的细胞外电子传输能力,从而提高BPV的效率和使用寿命,如图10 所示.该微生物组由能够将光能储存在D-乳酸的蓝藻和能够高效利用D-乳酸产电的希瓦菌组成,其中D-乳酸是两种微生物间的能量载体.蓝藻能够吸收光能并固定CO2 来合成能量载体D-乳酸,希瓦菌氧化D-乳酸进行产电,由此形成一条从光子到D-乳酸再到电能的定向电子流,完成从光能到化学能再到电能的能量转化过程.研究结果表明,该双菌生物光伏系统可以实现稳定、高效的功率输出,功率密度最高可达150 mW/m2 ,比普通单菌光伏系统高出10倍以上.该生物光伏技术可以使具有介质补充功能的装置稳定运行40 d以上,平均功率密度为135 mW/m2 ,在产电时长和功率输出方面均达到了较高水平.说明具有定向电子流的微生物组可以促进电子从光合细胞输出,并获得高效和持久的功率输出. ...
1
... 蓝藻是光合作用效率最高的生物之一,它们是原核生物,能量转换在类囊体膜中进行[43 ] .为了研究藻类用于制备生物光伏器件的能力,Ng等[44 ] 在氧化铟锡阳极和还原氧化石墨烯阳极上分别制备了长聚球藻的藻类生物膜,用于在光照和黑暗条件下研究藻类生物光伏器件.研究证明,与以氧化铟锡为阳极的生物光伏器件相比,以还原氧化石墨烯为阳极的生物光伏器件具有更高的功率输出,其功率密度最高可达0.538 mW/m2 ±0.014 mW/m2 ,说明了藻类生物光伏电池在光照下具有产生生物电的能力.Zhu等[45 ] 开发了一种具有定向电子流的以D-乳酸为中介的微生物组生物光伏技术,来提高光蓝藻的细胞外电子传输能力,从而提高BPV的效率和使用寿命,如图10 所示.该微生物组由能够将光能储存在D-乳酸的蓝藻和能够高效利用D-乳酸产电的希瓦菌组成,其中D-乳酸是两种微生物间的能量载体.蓝藻能够吸收光能并固定CO2 来合成能量载体D-乳酸,希瓦菌氧化D-乳酸进行产电,由此形成一条从光子到D-乳酸再到电能的定向电子流,完成从光能到化学能再到电能的能量转化过程.研究结果表明,该双菌生物光伏系统可以实现稳定、高效的功率输出,功率密度最高可达150 mW/m2 ,比普通单菌光伏系统高出10倍以上.该生物光伏技术可以使具有介质补充功能的装置稳定运行40 d以上,平均功率密度为135 mW/m2 ,在产电时长和功率输出方面均达到了较高水平.说明具有定向电子流的微生物组可以促进电子从光合细胞输出,并获得高效和持久的功率输出. ...
1
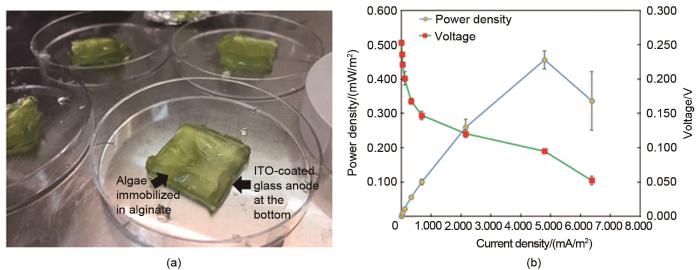
... Thong等[46 ] 分析了辐照度对藻类生物光伏装置输出功率的影响,确定了在氧化铟锡阳极上悬浮培养的小球藻和由海藻酸盐固定培养的小球藻[图11 (a)]组成的生物光伏装置的有效辐照度范围.小球藻固定化培养物是通过将小球藻细胞固定在2%的海藻酸钠溶液中制成.藻类生物光伏装置在4种不同的辐照度水平[30 μmol/(m2 · s)、90 μmol/(m2 · s)、150 μmol/(m2 · s)和210 μmol/(m2 · s)下]照射.实验结果显示,固定培养的小球藻光伏装置在150 μmol/(m2 · s)的辐照度下可产生0.456 mW/m2 的最大功率密度[图11 (b)],在210 μmol/(m2 · s)的辐照度下,由于光抑制,产生了低功率密度.此外,与悬浮培养相比,固定化培养的藻类生物光伏装置功率输出高32%.该团队认为这是因为海藻酸盐凝胶基质可以保护海藻细胞免受不利条件的影响,为细胞生长提供了有利的环境.此外,海藻酸盐中的细胞固定化导致海藻细胞在基质的基层上有了更大的沉积,这使穿过细胞与阳极界面的电子数目增加,导致功率输出密度增加. ...
1
... 据估计,1%的太阳能转化为电能所产生的能量是目前全世界所使用的总能量的10倍[47 ] .所以,对太阳能的有效利用一定是未来社会发展的大趋势,生物光伏系统就可以依托生物中间酶的高活性、植物的高效转化路径选择等助力这种趋势的发展.此外,生物光伏系统相对于传统光伏装置有一种独特的优势,即光敏组件由生物体组成,能够自我修复和繁殖.这可以减少设备的维护成本,并且节能环保.此外,类光合作用发供电技术主要依靠光营养微生物进行发供电,其产电过程是一个纯粹的生理过程,能够高效转化太阳能,产生稳定的生物能和电能,不会对环境造成污染,从而获得经济和环境效益的双提升;生物光伏技术中光敏组件由生物体组装和维护,能够进行自我修复和繁殖,降低设备维护成本,节能环保[48 ] ;可以通过对电极的设计和光照强度的调节提高产电性能,灵活性强;藻类等微生物还可吸收二氧化碳,具有碳中性的优点[49 ] . ...
1
... 据估计,1%的太阳能转化为电能所产生的能量是目前全世界所使用的总能量的10倍[47 ] .所以,对太阳能的有效利用一定是未来社会发展的大趋势,生物光伏系统就可以依托生物中间酶的高活性、植物的高效转化路径选择等助力这种趋势的发展.此外,生物光伏系统相对于传统光伏装置有一种独特的优势,即光敏组件由生物体组成,能够自我修复和繁殖.这可以减少设备的维护成本,并且节能环保.此外,类光合作用发供电技术主要依靠光营养微生物进行发供电,其产电过程是一个纯粹的生理过程,能够高效转化太阳能,产生稳定的生物能和电能,不会对环境造成污染,从而获得经济和环境效益的双提升;生物光伏技术中光敏组件由生物体组装和维护,能够进行自我修复和繁殖,降低设备维护成本,节能环保[48 ] ;可以通过对电极的设计和光照强度的调节提高产电性能,灵活性强;藻类等微生物还可吸收二氧化碳,具有碳中性的优点[49 ] . ...
1
... 据估计,1%的太阳能转化为电能所产生的能量是目前全世界所使用的总能量的10倍[47 ] .所以,对太阳能的有效利用一定是未来社会发展的大趋势,生物光伏系统就可以依托生物中间酶的高活性、植物的高效转化路径选择等助力这种趋势的发展.此外,生物光伏系统相对于传统光伏装置有一种独特的优势,即光敏组件由生物体组成,能够自我修复和繁殖.这可以减少设备的维护成本,并且节能环保.此外,类光合作用发供电技术主要依靠光营养微生物进行发供电,其产电过程是一个纯粹的生理过程,能够高效转化太阳能,产生稳定的生物能和电能,不会对环境造成污染,从而获得经济和环境效益的双提升;生物光伏技术中光敏组件由生物体组装和维护,能够进行自我修复和繁殖,降低设备维护成本,节能环保[48 ] ;可以通过对电极的设计和光照强度的调节提高产电性能,灵活性强;藻类等微生物还可吸收二氧化碳,具有碳中性的优点[49 ] . ...
1
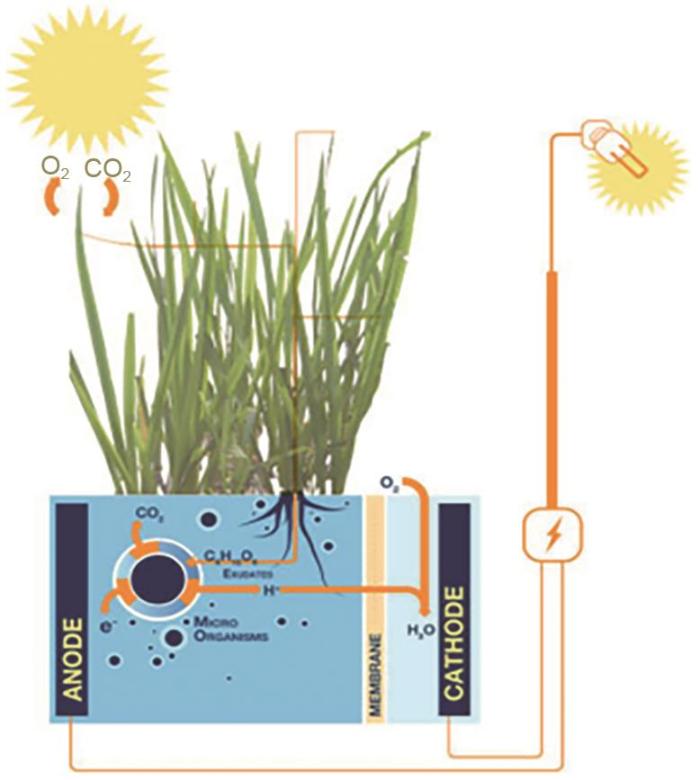
... 微生物燃料电池(MFC)是一种以微生物作为催化剂,将有机物中的化学能转化成电能的装置[50 ] .植物根系可以通过分泌物和一些相关过程将输入到根部的有机、无机化合物释放到周围土壤,形成根际沉积[51 ] ,这些有机沉积物为根际微生物的生命活动提供了有利条件.在根际沉积的土壤中应用MFC技术设计的装置称为沉积物微生物燃料电池(SMFC),这里的植物大多为水生植物,如水稻等.其工作原理如图12 所示,阳极材料放置于沉积物中,阴极材料多放置于富氧的水相中,阴极和阳极之间通过导线和电阻相连接,土壤中的有机物在阳极区附近被厌氧微生物氧化分解,产生的电子通过细胞膜传递到阳极,再经过外电路到达阴极,与阴极区中的氧气和从阳极区传递来的质子结合生成水,从而产生电能输出[52 ] .植物微生物燃料电池(PMFC)是指将植物与SMFC技术相结合,利用植物的光合作用,为产电微生物提供有机物,产电微生物在生长的同时产生电流,从而把太阳能转化为电能.对于该项技术,近年来取得的主要研究成果综述如下. ...
1
... 微生物燃料电池(MFC)是一种以微生物作为催化剂,将有机物中的化学能转化成电能的装置[50 ] .植物根系可以通过分泌物和一些相关过程将输入到根部的有机、无机化合物释放到周围土壤,形成根际沉积[51 ] ,这些有机沉积物为根际微生物的生命活动提供了有利条件.在根际沉积的土壤中应用MFC技术设计的装置称为沉积物微生物燃料电池(SMFC),这里的植物大多为水生植物,如水稻等.其工作原理如图12 所示,阳极材料放置于沉积物中,阴极材料多放置于富氧的水相中,阴极和阳极之间通过导线和电阻相连接,土壤中的有机物在阳极区附近被厌氧微生物氧化分解,产生的电子通过细胞膜传递到阳极,再经过外电路到达阴极,与阴极区中的氧气和从阳极区传递来的质子结合生成水,从而产生电能输出[52 ] .植物微生物燃料电池(PMFC)是指将植物与SMFC技术相结合,利用植物的光合作用,为产电微生物提供有机物,产电微生物在生长的同时产生电流,从而把太阳能转化为电能.对于该项技术,近年来取得的主要研究成果综述如下. ...
1
... 微生物燃料电池(MFC)是一种以微生物作为催化剂,将有机物中的化学能转化成电能的装置[50 ] .植物根系可以通过分泌物和一些相关过程将输入到根部的有机、无机化合物释放到周围土壤,形成根际沉积[51 ] ,这些有机沉积物为根际微生物的生命活动提供了有利条件.在根际沉积的土壤中应用MFC技术设计的装置称为沉积物微生物燃料电池(SMFC),这里的植物大多为水生植物,如水稻等.其工作原理如图12 所示,阳极材料放置于沉积物中,阴极材料多放置于富氧的水相中,阴极和阳极之间通过导线和电阻相连接,土壤中的有机物在阳极区附近被厌氧微生物氧化分解,产生的电子通过细胞膜传递到阳极,再经过外电路到达阴极,与阴极区中的氧气和从阳极区传递来的质子结合生成水,从而产生电能输出[52 ] .植物微生物燃料电池(PMFC)是指将植物与SMFC技术相结合,利用植物的光合作用,为产电微生物提供有机物,产电微生物在生长的同时产生电流,从而把太阳能转化为电能.对于该项技术,近年来取得的主要研究成果综述如下. ...
1
... 微生物燃料电池(MFC)是一种以微生物作为催化剂,将有机物中的化学能转化成电能的装置[50 ] .植物根系可以通过分泌物和一些相关过程将输入到根部的有机、无机化合物释放到周围土壤,形成根际沉积[51 ] ,这些有机沉积物为根际微生物的生命活动提供了有利条件.在根际沉积的土壤中应用MFC技术设计的装置称为沉积物微生物燃料电池(SMFC),这里的植物大多为水生植物,如水稻等.其工作原理如图12 所示,阳极材料放置于沉积物中,阴极材料多放置于富氧的水相中,阴极和阳极之间通过导线和电阻相连接,土壤中的有机物在阳极区附近被厌氧微生物氧化分解,产生的电子通过细胞膜传递到阳极,再经过外电路到达阴极,与阴极区中的氧气和从阳极区传递来的质子结合生成水,从而产生电能输出[52 ] .植物微生物燃料电池(PMFC)是指将植物与SMFC技术相结合,利用植物的光合作用,为产电微生物提供有机物,产电微生物在生长的同时产生电流,从而把太阳能转化为电能.对于该项技术,近年来取得的主要研究成果综述如下. ...
2
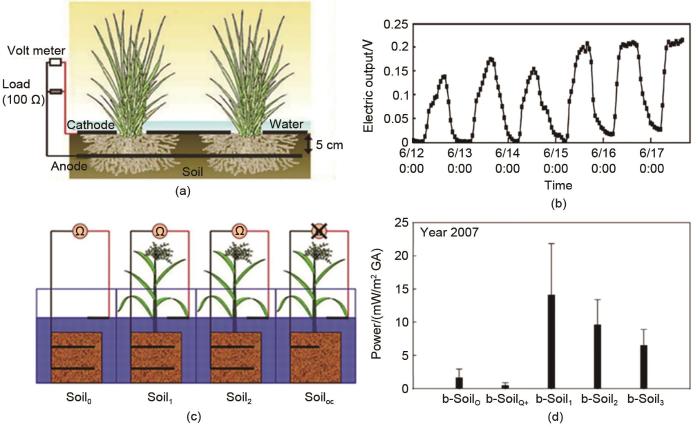
... Kaku等[53 ] 以石墨毡为电极,水稻为模型植物,设计了单室结构的植物微生物燃料电池,检测稻田中的发电情况,如图13 (a)所示.实验结果显示其发电量可达6 mW/m2 .在白天对水稻植株进行遮光处理后,其电压输出有所下降.此外,电压输出还表现出昼夜节律振荡,如图13 (b)所示,证明电压输出跟阳光有关.单室结构的植物微生物燃料电池的特点是结构简单,易于从自然环境中制备.类似地,Schamphelaire等[54 ] 用水稻制备了植物微生物燃料电池,研究在不同对照条件下燃料电池的发电情况.图13 (c)为燃料电池的示意图,4个独立隔间中可以选择种植或不种植植物,每个隔间都有一种供植物生根的支持基底,它包含一个或多个阳极,支持基底可以是常规土壤、蛭石等.在以土壤为基底的情况下,上述系统分别称为soil0 (闭路控制反应器,不含植物)、soil1 (闭路反应器,含植物)、soil2 (闭路反应器,含植物)和soiloc (开路反应器,含植物).研究发现,在有活植物存在时,微生物燃料电池的电能输出高出7倍,最大发电量可达33 mW/m2 ,证明了植物的存在可以增加发电量.图13 (d)为1年后进行的重复实验(b-soil),植物微生物燃料电池的输出功率密度仍远高于不含植物的微生物燃料电池. ...
... Summary of plant microbial fuel cell power generation performance parameters
Table 2 植物/培养基底 电子受体 最大功率密度/(mW/m2 ) 最大输出电压/mV 平均电流密度/(mA/m2 ) 参考文献 水稻/土壤 氧气 6.00 129 — [53 ] 水稻/土壤 铁氰化物 33.00 — 101~139 [54 ] 水稻/蛭石 铁氰化物 29.00 — 35~53 [54 ] 水稻/石墨 铁氰化物 17.90 — — [54 ] 水甜茅/石墨 氧气 67.00 253 — [55 ] 大米草/石墨 氧气 79.00 — 89~193 [60 ] 大米草/石墨 铁氰化物 100.00 — 111~279 [60 ] 野古草/石墨 铁氰化物 22.00 — 12~31 [65 ] 芦苇/石墨 氧气 26.78 320 19~53 [59 ]
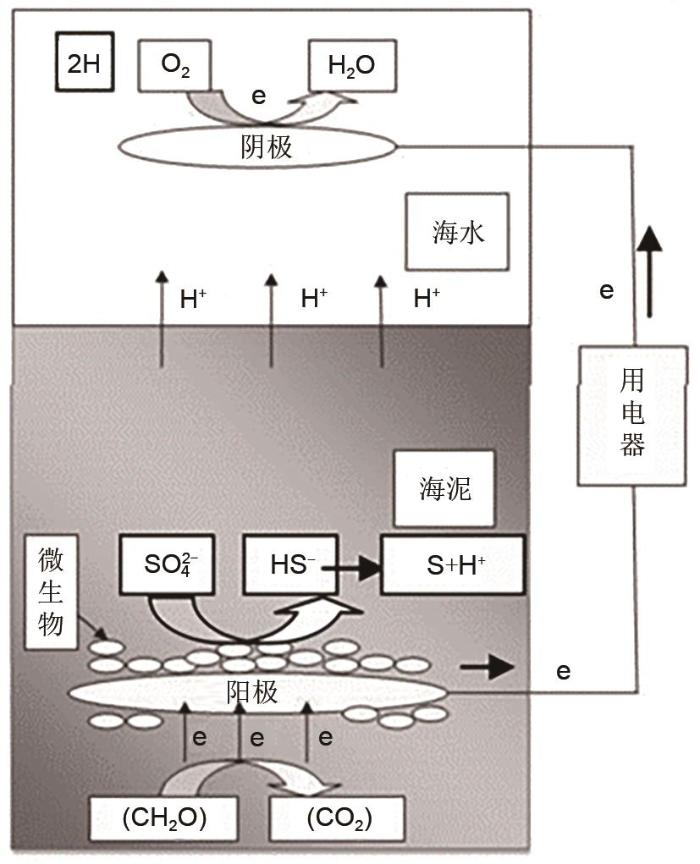
除了利用土壤和石墨颗粒等传统培养基底供产电微生物生长和发电,一部分燃料电池以海底沉积物(海泥)作为能量来源,通过微生物代谢有机物而产生电能,称为海底沉积物生物燃料电池(简称海底生物燃料电池).海底生物燃料电池主要利用天然的海泥层作为阳极导电介质,以有机物和无机物作为燃料及电子来源,以海水层作为阴极的导电介质,海水溶解氧作为氧化剂,消耗电子[66 ] .其工作原理如图16 所示,在海泥中埋置导电碳材料作为电极(阳极),细菌附着在阳极表面,将自身代谢产生的电子由外电路传输到海水中的阴极,质子通过海底沉积层/海水界面扩散到阴极,在阴极表面发生海水溶解氧的还原反应生成水,从而构成电池回路,产生电流[67 ] . ...
4
... Kaku等[53 ] 以石墨毡为电极,水稻为模型植物,设计了单室结构的植物微生物燃料电池,检测稻田中的发电情况,如图13 (a)所示.实验结果显示其发电量可达6 mW/m2 .在白天对水稻植株进行遮光处理后,其电压输出有所下降.此外,电压输出还表现出昼夜节律振荡,如图13 (b)所示,证明电压输出跟阳光有关.单室结构的植物微生物燃料电池的特点是结构简单,易于从自然环境中制备.类似地,Schamphelaire等[54 ] 用水稻制备了植物微生物燃料电池,研究在不同对照条件下燃料电池的发电情况.图13 (c)为燃料电池的示意图,4个独立隔间中可以选择种植或不种植植物,每个隔间都有一种供植物生根的支持基底,它包含一个或多个阳极,支持基底可以是常规土壤、蛭石等.在以土壤为基底的情况下,上述系统分别称为soil0 (闭路控制反应器,不含植物)、soil1 (闭路反应器,含植物)、soil2 (闭路反应器,含植物)和soiloc (开路反应器,含植物).研究发现,在有活植物存在时,微生物燃料电池的电能输出高出7倍,最大发电量可达33 mW/m2 ,证明了植物的存在可以增加发电量.图13 (d)为1年后进行的重复实验(b-soil),植物微生物燃料电池的输出功率密度仍远高于不含植物的微生物燃料电池. ...
... Summary of plant microbial fuel cell power generation performance parameters
Table 2 植物/培养基底 电子受体 最大功率密度/(mW/m2 ) 最大输出电压/mV 平均电流密度/(mA/m2 ) 参考文献 水稻/土壤 氧气 6.00 129 — [53 ] 水稻/土壤 铁氰化物 33.00 — 101~139 [54 ] 水稻/蛭石 铁氰化物 29.00 — 35~53 [54 ] 水稻/石墨 铁氰化物 17.90 — — [54 ] 水甜茅/石墨 氧气 67.00 253 — [55 ] 大米草/石墨 氧气 79.00 — 89~193 [60 ] 大米草/石墨 铁氰化物 100.00 — 111~279 [60 ] 野古草/石墨 铁氰化物 22.00 — 12~31 [65 ] 芦苇/石墨 氧气 26.78 320 19~53 [59 ]
除了利用土壤和石墨颗粒等传统培养基底供产电微生物生长和发电,一部分燃料电池以海底沉积物(海泥)作为能量来源,通过微生物代谢有机物而产生电能,称为海底沉积物生物燃料电池(简称海底生物燃料电池).海底生物燃料电池主要利用天然的海泥层作为阳极导电介质,以有机物和无机物作为燃料及电子来源,以海水层作为阴极的导电介质,海水溶解氧作为氧化剂,消耗电子[66 ] .其工作原理如图16 所示,在海泥中埋置导电碳材料作为电极(阳极),细菌附着在阳极表面,将自身代谢产生的电子由外电路传输到海水中的阴极,质子通过海底沉积层/海水界面扩散到阴极,在阴极表面发生海水溶解氧的还原反应生成水,从而构成电池回路,产生电流[67 ] . ...
... [
54 ]
水稻/石墨 铁氰化物 17.90 — — [54 ] 水甜茅/石墨 氧气 67.00 253 — [55 ] 大米草/石墨 氧气 79.00 — 89~193 [60 ] 大米草/石墨 铁氰化物 100.00 — 111~279 [60 ] 野古草/石墨 铁氰化物 22.00 — 12~31 [65 ] 芦苇/石墨 氧气 26.78 320 19~53 [59 ] 除了利用土壤和石墨颗粒等传统培养基底供产电微生物生长和发电,一部分燃料电池以海底沉积物(海泥)作为能量来源,通过微生物代谢有机物而产生电能,称为海底沉积物生物燃料电池(简称海底生物燃料电池).海底生物燃料电池主要利用天然的海泥层作为阳极导电介质,以有机物和无机物作为燃料及电子来源,以海水层作为阴极的导电介质,海水溶解氧作为氧化剂,消耗电子[66 ] .其工作原理如图16 所示,在海泥中埋置导电碳材料作为电极(阳极),细菌附着在阳极表面,将自身代谢产生的电子由外电路传输到海水中的阴极,质子通过海底沉积层/海水界面扩散到阴极,在阴极表面发生海水溶解氧的还原反应生成水,从而构成电池回路,产生电流[67 ] . ...
... [
54 ]
水甜茅/石墨 氧气 67.00 253 — [55 ] 大米草/石墨 氧气 79.00 — 89~193 [60 ] 大米草/石墨 铁氰化物 100.00 — 111~279 [60 ] 野古草/石墨 铁氰化物 22.00 — 12~31 [65 ] 芦苇/石墨 氧气 26.78 320 19~53 [59 ] 除了利用土壤和石墨颗粒等传统培养基底供产电微生物生长和发电,一部分燃料电池以海底沉积物(海泥)作为能量来源,通过微生物代谢有机物而产生电能,称为海底沉积物生物燃料电池(简称海底生物燃料电池).海底生物燃料电池主要利用天然的海泥层作为阳极导电介质,以有机物和无机物作为燃料及电子来源,以海水层作为阴极的导电介质,海水溶解氧作为氧化剂,消耗电子[66 ] .其工作原理如图16 所示,在海泥中埋置导电碳材料作为电极(阳极),细菌附着在阳极表面,将自身代谢产生的电子由外电路传输到海水中的阴极,质子通过海底沉积层/海水界面扩散到阴极,在阴极表面发生海水溶解氧的还原反应生成水,从而构成电池回路,产生电流[67 ] . ...
3
... 除常用的水稻植物微生物燃料电池,Strik团队[55 ] 利用水甜茅作为模型植物设计了双室结构的燃料电池,在阳极表面得到了67 mW/m2 的最大发电量.该植物微生物燃料电池具有以下特点:①生物能源的无损、原位采集;②可以在湿地和贫瘠的土壤中实施,不与食物或一些传统生物能源产生竞争;③在欧洲,估计年潜在发电量可达每公顷5800 kW· h;④碳中和、无排放燃烧.双室结构的植物微生物燃料电池的电量输出一般较高,且阴阳极室可以选择不同的反应液,但需要使用质子交换膜,增加成本,膜的长期使用还会造成堵塞,使电池内阻增加[56 ] .另外,Castresana等[57 ] 选择苔藓作为模型植物,报道了一种无膜光驱动植物微生物燃料电池,并研究了它从苔藓中获取电能的能力.图14 为植物微生物燃料电池和阴极沉积材料的模型.阴极使用CuO-Cu2 O作光催化剂,利用Nafion涂层防止光催化剂浸出.在辐照下,光生电子与阴极/土壤界面的电子受体发生反应.Cux x
... 依靠微生物氧化分解根际沉积物产电的传统植物微生物燃料电池的发电能力一般会受环境因素和植物种类的影响.鉴于此,Lu等[61 ] 用利发财树和银白杨,设计了一种可以直接从植物茎中产生连续生物电的燃料电池,如图15 所示.与传统的植物微生物燃料电池相比,该新型电池受光循环的影响较小,产生的电流更为稳定和连续,启动时间也短得多.这是因为每日的光循环会显著影响植物的光合作用和根系沉积物的产生[55 ,62 ] ,而植物茎的基质供应在短时间内较少受到阳光的影响,从而产生强且连续的生物电.此外,相比于与发财树,与银白杨结合的微生物燃料电池具有更高的功率输出,其最大功率密度分别为7.61 mW/m2 . ...
... Summary of plant microbial fuel cell power generation performance parameters
Table 2 植物/培养基底 电子受体 最大功率密度/(mW/m2 ) 最大输出电压/mV 平均电流密度/(mA/m2 ) 参考文献 水稻/土壤 氧气 6.00 129 — [53 ] 水稻/土壤 铁氰化物 33.00 — 101~139 [54 ] 水稻/蛭石 铁氰化物 29.00 — 35~53 [54 ] 水稻/石墨 铁氰化物 17.90 — — [54 ] 水甜茅/石墨 氧气 67.00 253 — [55 ] 大米草/石墨 氧气 79.00 — 89~193 [60 ] 大米草/石墨 铁氰化物 100.00 — 111~279 [60 ] 野古草/石墨 铁氰化物 22.00 — 12~31 [65 ] 芦苇/石墨 氧气 26.78 320 19~53 [59 ]
除了利用土壤和石墨颗粒等传统培养基底供产电微生物生长和发电,一部分燃料电池以海底沉积物(海泥)作为能量来源,通过微生物代谢有机物而产生电能,称为海底沉积物生物燃料电池(简称海底生物燃料电池).海底生物燃料电池主要利用天然的海泥层作为阳极导电介质,以有机物和无机物作为燃料及电子来源,以海水层作为阴极的导电介质,海水溶解氧作为氧化剂,消耗电子[66 ] .其工作原理如图16 所示,在海泥中埋置导电碳材料作为电极(阳极),细菌附着在阳极表面,将自身代谢产生的电子由外电路传输到海水中的阴极,质子通过海底沉积层/海水界面扩散到阴极,在阴极表面发生海水溶解氧的还原反应生成水,从而构成电池回路,产生电流[67 ] . ...
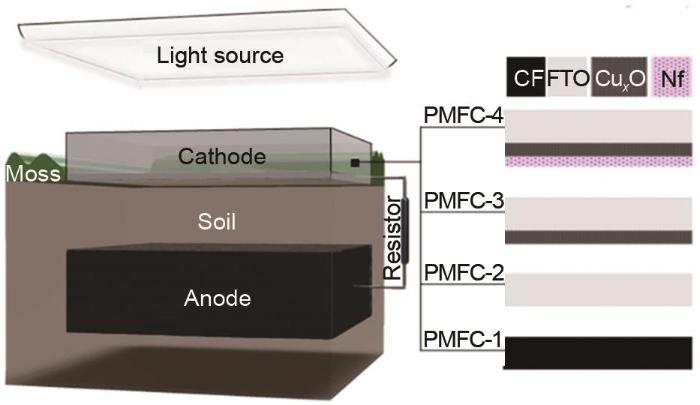
1
... 除常用的水稻植物微生物燃料电池,Strik团队[55 ] 利用水甜茅作为模型植物设计了双室结构的燃料电池,在阳极表面得到了67 mW/m2 的最大发电量.该植物微生物燃料电池具有以下特点:①生物能源的无损、原位采集;②可以在湿地和贫瘠的土壤中实施,不与食物或一些传统生物能源产生竞争;③在欧洲,估计年潜在发电量可达每公顷5800 kW· h;④碳中和、无排放燃烧.双室结构的植物微生物燃料电池的电量输出一般较高,且阴阳极室可以选择不同的反应液,但需要使用质子交换膜,增加成本,膜的长期使用还会造成堵塞,使电池内阻增加[56 ] .另外,Castresana等[57 ] 选择苔藓作为模型植物,报道了一种无膜光驱动植物微生物燃料电池,并研究了它从苔藓中获取电能的能力.图14 为植物微生物燃料电池和阴极沉积材料的模型.阴极使用CuO-Cu2 O作光催化剂,利用Nafion涂层防止光催化剂浸出.在辐照下,光生电子与阴极/土壤界面的电子受体发生反应.Cux x
1
... 除常用的水稻植物微生物燃料电池,Strik团队[55 ] 利用水甜茅作为模型植物设计了双室结构的燃料电池,在阳极表面得到了67 mW/m2 的最大发电量.该植物微生物燃料电池具有以下特点:①生物能源的无损、原位采集;②可以在湿地和贫瘠的土壤中实施,不与食物或一些传统生物能源产生竞争;③在欧洲,估计年潜在发电量可达每公顷5800 kW· h;④碳中和、无排放燃烧.双室结构的植物微生物燃料电池的电量输出一般较高,且阴阳极室可以选择不同的反应液,但需要使用质子交换膜,增加成本,膜的长期使用还会造成堵塞,使电池内阻增加[56 ] .另外,Castresana等[57 ] 选择苔藓作为模型植物,报道了一种无膜光驱动植物微生物燃料电池,并研究了它从苔藓中获取电能的能力.图14 为植物微生物燃料电池和阴极沉积材料的模型.阴极使用CuO-Cu2 O作光催化剂,利用Nafion涂层防止光催化剂浸出.在辐照下,光生电子与阴极/土壤界面的电子受体发生反应.Cux x
1
... 除常用的水稻植物微生物燃料电池,Strik团队[55 ] 利用水甜茅作为模型植物设计了双室结构的燃料电池,在阳极表面得到了67 mW/m2 的最大发电量.该植物微生物燃料电池具有以下特点:①生物能源的无损、原位采集;②可以在湿地和贫瘠的土壤中实施,不与食物或一些传统生物能源产生竞争;③在欧洲,估计年潜在发电量可达每公顷5800 kW· h;④碳中和、无排放燃烧.双室结构的植物微生物燃料电池的电量输出一般较高,且阴阳极室可以选择不同的反应液,但需要使用质子交换膜,增加成本,膜的长期使用还会造成堵塞,使电池内阻增加[56 ] .另外,Castresana等[57 ] 选择苔藓作为模型植物,报道了一种无膜光驱动植物微生物燃料电池,并研究了它从苔藓中获取电能的能力.图14 为植物微生物燃料电池和阴极沉积材料的模型.阴极使用CuO-Cu2 O作光催化剂,利用Nafion涂层防止光催化剂浸出.在辐照下,光生电子与阴极/土壤界面的电子受体发生反应.Cux x
1
... 在植物微生物燃料电池工作时,会发生一系列物理和生物化学过程,植物的培养基底可能会影响其电能输出.基底应为微生物的健壮生长、代谢活性、降解有机物等提供合适的条件,并能够将电子转移到阳极[58 ] .为了验证植物微生物燃料电池在人工湿地中的实施情况,Salinas-Juárez等[59 ] 以芦苇为模型植物,石墨颗粒和火山渣作为植物的支持基底,制备了植物微生物燃料电池,研究其产电能力.结果表明,两者都有潜力作为植物和微生物的支持基底,在石墨颗粒基底中可以产生的最大输出功率为26.78 mW/m2 .Timmers等[60 ] 利用盐沼物种大米草设计了植物微生物燃料电池并对其供电性能进行研究,该燃料电池系统的培养基底为石墨颗粒,电极为石墨毡.在实验中,该团队在阴极加入铁氰化物作为电子受体,结果使供电性能提高,最大输出功率可达100 mW/m2 ,而阴极未加入铁氰化物时(氧气作为电子受体),最大输出功率密度为79 mW/m2 .这是由于阴极氧还原动力学缓慢,阴极过电位是氧还原期间的主要电位损失,而铁氰化物的还原改善了阴极的动力学,导致阳极电位增加,使电性能提高.该研究证明了阴极的电子受体会影响植物微生物燃料电池的供电性能,同时也验证了在盐沼中利用植物微生物燃料电池生产电能的可能性. ...
2
... 在植物微生物燃料电池工作时,会发生一系列物理和生物化学过程,植物的培养基底可能会影响其电能输出.基底应为微生物的健壮生长、代谢活性、降解有机物等提供合适的条件,并能够将电子转移到阳极[58 ] .为了验证植物微生物燃料电池在人工湿地中的实施情况,Salinas-Juárez等[59 ] 以芦苇为模型植物,石墨颗粒和火山渣作为植物的支持基底,制备了植物微生物燃料电池,研究其产电能力.结果表明,两者都有潜力作为植物和微生物的支持基底,在石墨颗粒基底中可以产生的最大输出功率为26.78 mW/m2 .Timmers等[60 ] 利用盐沼物种大米草设计了植物微生物燃料电池并对其供电性能进行研究,该燃料电池系统的培养基底为石墨颗粒,电极为石墨毡.在实验中,该团队在阴极加入铁氰化物作为电子受体,结果使供电性能提高,最大输出功率可达100 mW/m2 ,而阴极未加入铁氰化物时(氧气作为电子受体),最大输出功率密度为79 mW/m2 .这是由于阴极氧还原动力学缓慢,阴极过电位是氧还原期间的主要电位损失,而铁氰化物的还原改善了阴极的动力学,导致阳极电位增加,使电性能提高.该研究证明了阴极的电子受体会影响植物微生物燃料电池的供电性能,同时也验证了在盐沼中利用植物微生物燃料电池生产电能的可能性. ...
... Summary of plant microbial fuel cell power generation performance parameters
Table 2 植物/培养基底 电子受体 最大功率密度/(mW/m2 ) 最大输出电压/mV 平均电流密度/(mA/m2 ) 参考文献 水稻/土壤 氧气 6.00 129 — [53 ] 水稻/土壤 铁氰化物 33.00 — 101~139 [54 ] 水稻/蛭石 铁氰化物 29.00 — 35~53 [54 ] 水稻/石墨 铁氰化物 17.90 — — [54 ] 水甜茅/石墨 氧气 67.00 253 — [55 ] 大米草/石墨 氧气 79.00 — 89~193 [60 ] 大米草/石墨 铁氰化物 100.00 — 111~279 [60 ] 野古草/石墨 铁氰化物 22.00 — 12~31 [65 ] 芦苇/石墨 氧气 26.78 320 19~53 [59 ]
除了利用土壤和石墨颗粒等传统培养基底供产电微生物生长和发电,一部分燃料电池以海底沉积物(海泥)作为能量来源,通过微生物代谢有机物而产生电能,称为海底沉积物生物燃料电池(简称海底生物燃料电池).海底生物燃料电池主要利用天然的海泥层作为阳极导电介质,以有机物和无机物作为燃料及电子来源,以海水层作为阴极的导电介质,海水溶解氧作为氧化剂,消耗电子[66 ] .其工作原理如图16 所示,在海泥中埋置导电碳材料作为电极(阳极),细菌附着在阳极表面,将自身代谢产生的电子由外电路传输到海水中的阴极,质子通过海底沉积层/海水界面扩散到阴极,在阴极表面发生海水溶解氧的还原反应生成水,从而构成电池回路,产生电流[67 ] . ...
3
... 在植物微生物燃料电池工作时,会发生一系列物理和生物化学过程,植物的培养基底可能会影响其电能输出.基底应为微生物的健壮生长、代谢活性、降解有机物等提供合适的条件,并能够将电子转移到阳极[58 ] .为了验证植物微生物燃料电池在人工湿地中的实施情况,Salinas-Juárez等[59 ] 以芦苇为模型植物,石墨颗粒和火山渣作为植物的支持基底,制备了植物微生物燃料电池,研究其产电能力.结果表明,两者都有潜力作为植物和微生物的支持基底,在石墨颗粒基底中可以产生的最大输出功率为26.78 mW/m2 .Timmers等[60 ] 利用盐沼物种大米草设计了植物微生物燃料电池并对其供电性能进行研究,该燃料电池系统的培养基底为石墨颗粒,电极为石墨毡.在实验中,该团队在阴极加入铁氰化物作为电子受体,结果使供电性能提高,最大输出功率可达100 mW/m2 ,而阴极未加入铁氰化物时(氧气作为电子受体),最大输出功率密度为79 mW/m2 .这是由于阴极氧还原动力学缓慢,阴极过电位是氧还原期间的主要电位损失,而铁氰化物的还原改善了阴极的动力学,导致阳极电位增加,使电性能提高.该研究证明了阴极的电子受体会影响植物微生物燃料电池的供电性能,同时也验证了在盐沼中利用植物微生物燃料电池生产电能的可能性. ...
... Summary of plant microbial fuel cell power generation performance parameters
Table 2 植物/培养基底 电子受体 最大功率密度/(mW/m2 ) 最大输出电压/mV 平均电流密度/(mA/m2 ) 参考文献 水稻/土壤 氧气 6.00 129 — [53 ] 水稻/土壤 铁氰化物 33.00 — 101~139 [54 ] 水稻/蛭石 铁氰化物 29.00 — 35~53 [54 ] 水稻/石墨 铁氰化物 17.90 — — [54 ] 水甜茅/石墨 氧气 67.00 253 — [55 ] 大米草/石墨 氧气 79.00 — 89~193 [60 ] 大米草/石墨 铁氰化物 100.00 — 111~279 [60 ] 野古草/石墨 铁氰化物 22.00 — 12~31 [65 ] 芦苇/石墨 氧气 26.78 320 19~53 [59 ]
除了利用土壤和石墨颗粒等传统培养基底供产电微生物生长和发电,一部分燃料电池以海底沉积物(海泥)作为能量来源,通过微生物代谢有机物而产生电能,称为海底沉积物生物燃料电池(简称海底生物燃料电池).海底生物燃料电池主要利用天然的海泥层作为阳极导电介质,以有机物和无机物作为燃料及电子来源,以海水层作为阴极的导电介质,海水溶解氧作为氧化剂,消耗电子[66 ] .其工作原理如图16 所示,在海泥中埋置导电碳材料作为电极(阳极),细菌附着在阳极表面,将自身代谢产生的电子由外电路传输到海水中的阴极,质子通过海底沉积层/海水界面扩散到阴极,在阴极表面发生海水溶解氧的还原反应生成水,从而构成电池回路,产生电流[67 ] . ...
... [
60 ]
野古草/石墨 铁氰化物 22.00 — 12~31 [65 ] 芦苇/石墨 氧气 26.78 320 19~53 [59 ] 除了利用土壤和石墨颗粒等传统培养基底供产电微生物生长和发电,一部分燃料电池以海底沉积物(海泥)作为能量来源,通过微生物代谢有机物而产生电能,称为海底沉积物生物燃料电池(简称海底生物燃料电池).海底生物燃料电池主要利用天然的海泥层作为阳极导电介质,以有机物和无机物作为燃料及电子来源,以海水层作为阴极的导电介质,海水溶解氧作为氧化剂,消耗电子[66 ] .其工作原理如图16 所示,在海泥中埋置导电碳材料作为电极(阳极),细菌附着在阳极表面,将自身代谢产生的电子由外电路传输到海水中的阴极,质子通过海底沉积层/海水界面扩散到阴极,在阴极表面发生海水溶解氧的还原反应生成水,从而构成电池回路,产生电流[67 ] . ...
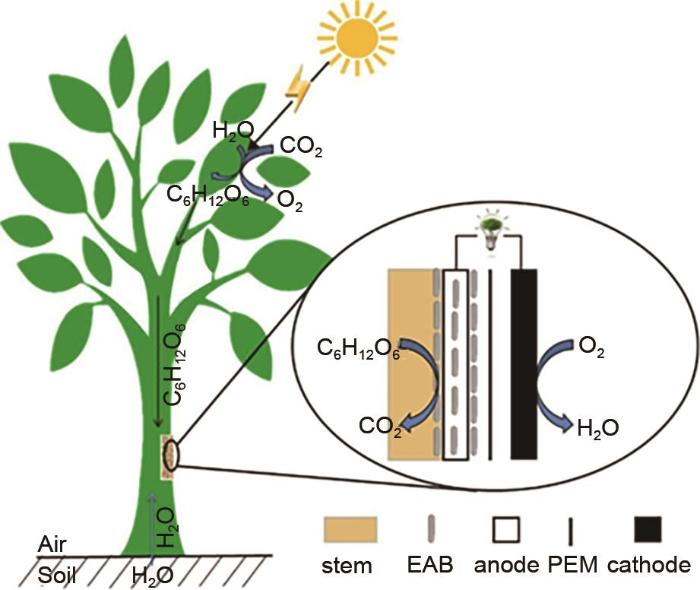
1
... 依靠微生物氧化分解根际沉积物产电的传统植物微生物燃料电池的发电能力一般会受环境因素和植物种类的影响.鉴于此,Lu等[61 ] 用利发财树和银白杨,设计了一种可以直接从植物茎中产生连续生物电的燃料电池,如图15 所示.与传统的植物微生物燃料电池相比,该新型电池受光循环的影响较小,产生的电流更为稳定和连续,启动时间也短得多.这是因为每日的光循环会显著影响植物的光合作用和根系沉积物的产生[55 ,62 ] ,而植物茎的基质供应在短时间内较少受到阳光的影响,从而产生强且连续的生物电.此外,相比于与发财树,与银白杨结合的微生物燃料电池具有更高的功率输出,其最大功率密度分别为7.61 mW/m2 . ...
1
... 依靠微生物氧化分解根际沉积物产电的传统植物微生物燃料电池的发电能力一般会受环境因素和植物种类的影响.鉴于此,Lu等[61 ] 用利发财树和银白杨,设计了一种可以直接从植物茎中产生连续生物电的燃料电池,如图15 所示.与传统的植物微生物燃料电池相比,该新型电池受光循环的影响较小,产生的电流更为稳定和连续,启动时间也短得多.这是因为每日的光循环会显著影响植物的光合作用和根系沉积物的产生[55 ,62 ] ,而植物茎的基质供应在短时间内较少受到阳光的影响,从而产生强且连续的生物电.此外,相比于与发财树,与银白杨结合的微生物燃料电池具有更高的功率输出,其最大功率密度分别为7.61 mW/m2 . ...
1
... 此外,通过对微生物菌群进行优选及组合培育,可以提高燃料电池的输出功率,用于高效供电系统的开发.混合菌落微生物燃料电池直接利用环境中的多种微生物作为催化剂产电,相较于单一菌种的微生物燃料电池,混合菌群电池启动省时、节约成本、环境兼容性好、可利用基质范围广,同时可以发挥菌群间的协同作用,增强微生物燃料电池运行的稳定性,提高系统的产电效率[63 ] .例如,Song等[64 ] 设计了一个由大肠杆菌、枯草芽孢杆菌和希瓦菌组成的微生物群落,用于发电.在该混合菌落中,大肠杆菌分解葡萄糖产生乳酸盐作为碳源和电子供体;枯草芽孢杆菌产生核黄素作为电子穿梭体,促进电子转移;希瓦菌作为电化学活性菌发电.反过来,希瓦菌将乳酸氧化成乙酸,乙酸作为碳源供给大肠杆菌和枯草芽孢杆菌.因此,3个菌种形成了一个“交叉喂养”的混合菌落,通过协效作用,提高了燃料电池的产电效率.此外,希瓦菌一般只能利用有限的碳源,如乳酸盐和丙酮酸盐,利用大肠杆菌分解葡萄糖产生乳酸盐,供给希瓦菌,丰富了希瓦菌可以利用的碳源.混合菌落微生物燃料电池为设计更稳定高效的、用于产电和环境修复的系统提供了新的方法. ...
1
... 此外,通过对微生物菌群进行优选及组合培育,可以提高燃料电池的输出功率,用于高效供电系统的开发.混合菌落微生物燃料电池直接利用环境中的多种微生物作为催化剂产电,相较于单一菌种的微生物燃料电池,混合菌群电池启动省时、节约成本、环境兼容性好、可利用基质范围广,同时可以发挥菌群间的协同作用,增强微生物燃料电池运行的稳定性,提高系统的产电效率[63 ] .例如,Song等[64 ] 设计了一个由大肠杆菌、枯草芽孢杆菌和希瓦菌组成的微生物群落,用于发电.在该混合菌落中,大肠杆菌分解葡萄糖产生乳酸盐作为碳源和电子供体;枯草芽孢杆菌产生核黄素作为电子穿梭体,促进电子转移;希瓦菌作为电化学活性菌发电.反过来,希瓦菌将乳酸氧化成乙酸,乙酸作为碳源供给大肠杆菌和枯草芽孢杆菌.因此,3个菌种形成了一个“交叉喂养”的混合菌落,通过协效作用,提高了燃料电池的产电效率.此外,希瓦菌一般只能利用有限的碳源,如乳酸盐和丙酮酸盐,利用大肠杆菌分解葡萄糖产生乳酸盐,供给希瓦菌,丰富了希瓦菌可以利用的碳源.混合菌落微生物燃料电池为设计更稳定高效的、用于产电和环境修复的系统提供了新的方法. ...
1
... 此外,通过对微生物菌群进行优选及组合培育,可以提高燃料电池的输出功率,用于高效供电系统的开发.混合菌落微生物燃料电池直接利用环境中的多种微生物作为催化剂产电,相较于单一菌种的微生物燃料电池,混合菌群电池启动省时、节约成本、环境兼容性好、可利用基质范围广,同时可以发挥菌群间的协同作用,增强微生物燃料电池运行的稳定性,提高系统的产电效率[63 ] .例如,Song等[64 ] 设计了一个由大肠杆菌、枯草芽孢杆菌和希瓦菌组成的微生物群落,用于发电.在该混合菌落中,大肠杆菌分解葡萄糖产生乳酸盐作为碳源和电子供体;枯草芽孢杆菌产生核黄素作为电子穿梭体,促进电子转移;希瓦菌作为电化学活性菌发电.反过来,希瓦菌将乳酸氧化成乙酸,乙酸作为碳源供给大肠杆菌和枯草芽孢杆菌.因此,3个菌种形成了一个“交叉喂养”的混合菌落,通过协效作用,提高了燃料电池的产电效率.此外,希瓦菌一般只能利用有限的碳源,如乳酸盐和丙酮酸盐,利用大肠杆菌分解葡萄糖产生乳酸盐,供给希瓦菌,丰富了希瓦菌可以利用的碳源.混合菌落微生物燃料电池为设计更稳定高效的、用于产电和环境修复的系统提供了新的方法. ...
1
... Summary of plant microbial fuel cell power generation performance parameters
Table 2 植物/培养基底 电子受体 最大功率密度/(mW/m2 ) 最大输出电压/mV 平均电流密度/(mA/m2 ) 参考文献 水稻/土壤 氧气 6.00 129 — [53 ] 水稻/土壤 铁氰化物 33.00 — 101~139 [54 ] 水稻/蛭石 铁氰化物 29.00 — 35~53 [54 ] 水稻/石墨 铁氰化物 17.90 — — [54 ] 水甜茅/石墨 氧气 67.00 253 — [55 ] 大米草/石墨 氧气 79.00 — 89~193 [60 ] 大米草/石墨 铁氰化物 100.00 — 111~279 [60 ] 野古草/石墨 铁氰化物 22.00 — 12~31 [65 ] 芦苇/石墨 氧气 26.78 320 19~53 [59 ]
除了利用土壤和石墨颗粒等传统培养基底供产电微生物生长和发电,一部分燃料电池以海底沉积物(海泥)作为能量来源,通过微生物代谢有机物而产生电能,称为海底沉积物生物燃料电池(简称海底生物燃料电池).海底生物燃料电池主要利用天然的海泥层作为阳极导电介质,以有机物和无机物作为燃料及电子来源,以海水层作为阴极的导电介质,海水溶解氧作为氧化剂,消耗电子[66 ] .其工作原理如图16 所示,在海泥中埋置导电碳材料作为电极(阳极),细菌附着在阳极表面,将自身代谢产生的电子由外电路传输到海水中的阴极,质子通过海底沉积层/海水界面扩散到阴极,在阴极表面发生海水溶解氧的还原反应生成水,从而构成电池回路,产生电流[67 ] . ...
1
... 除了利用土壤和石墨颗粒等传统培养基底供产电微生物生长和发电,一部分燃料电池以海底沉积物(海泥)作为能量来源,通过微生物代谢有机物而产生电能,称为海底沉积物生物燃料电池(简称海底生物燃料电池).海底生物燃料电池主要利用天然的海泥层作为阳极导电介质,以有机物和无机物作为燃料及电子来源,以海水层作为阴极的导电介质,海水溶解氧作为氧化剂,消耗电子[66 ] .其工作原理如图16 所示,在海泥中埋置导电碳材料作为电极(阳极),细菌附着在阳极表面,将自身代谢产生的电子由外电路传输到海水中的阴极,质子通过海底沉积层/海水界面扩散到阴极,在阴极表面发生海水溶解氧的还原反应生成水,从而构成电池回路,产生电流[67 ] . ...
1
... 除了利用土壤和石墨颗粒等传统培养基底供产电微生物生长和发电,一部分燃料电池以海底沉积物(海泥)作为能量来源,通过微生物代谢有机物而产生电能,称为海底沉积物生物燃料电池(简称海底生物燃料电池).海底生物燃料电池主要利用天然的海泥层作为阳极导电介质,以有机物和无机物作为燃料及电子来源,以海水层作为阴极的导电介质,海水溶解氧作为氧化剂,消耗电子[66 ] .其工作原理如图16 所示,在海泥中埋置导电碳材料作为电极(阳极),细菌附着在阳极表面,将自身代谢产生的电子由外电路传输到海水中的阴极,质子通过海底沉积层/海水界面扩散到阴极,在阴极表面发生海水溶解氧的还原反应生成水,从而构成电池回路,产生电流[67 ] . ...
1
... 除了利用土壤和石墨颗粒等传统培养基底供产电微生物生长和发电,一部分燃料电池以海底沉积物(海泥)作为能量来源,通过微生物代谢有机物而产生电能,称为海底沉积物生物燃料电池(简称海底生物燃料电池).海底生物燃料电池主要利用天然的海泥层作为阳极导电介质,以有机物和无机物作为燃料及电子来源,以海水层作为阴极的导电介质,海水溶解氧作为氧化剂,消耗电子[66 ] .其工作原理如图16 所示,在海泥中埋置导电碳材料作为电极(阳极),细菌附着在阳极表面,将自身代谢产生的电子由外电路传输到海水中的阴极,质子通过海底沉积层/海水界面扩散到阴极,在阴极表面发生海水溶解氧的还原反应生成水,从而构成电池回路,产生电流[67 ] . ...
1
... 一般而言,海底生物燃料电池的输出电压和输出功率较低,限制其广泛应用.因此,目前大多数研究集中于阳极改性上,这是提高电池电压和输出功率的有效手段.Prakash等[68 ] 研究了石墨板和碳布两种阳极材料经表面处理和表面涂层改性后的电性能.该团队制备了4种不同的改性阳极:酸处理碳布、聚苯胺涂覆碳布、酸处理后开槽的石墨板和聚苯胺涂覆普通石墨板.4种阳极均先使用25%的盐酸在40 ℃下加热处理20 min,目的是除去有机成分,同时在缺陷位置引入羧酸基团,有助于在阳极的羧基和苯胺的氨基之间形成氢键.开槽处理目的是在保持阳极尺寸不变的同时增加表面积和表面粗糙度,获得更好的生物膜黏附性,凹槽尺寸为宽1 mm、高0.8 mm.通过实验,得到了不同改性阳极的最大输出功率密度:聚苯胺涂覆碳布为11.14 mW/m2 ,酸处理碳布为5.75 mW/m2 ,聚苯胺涂覆普通石墨板为5.52 mW/m2 ,酸处理后开槽的石墨板为2.86 mW/m2 .研究也证明了聚苯胺涂覆的阳极材料适用于腐蚀性海洋环境,并支持生物膜生长,提高发电量.Fu等[69 ] 报道了一种用电解沉积法制备的铁/氧化铁改性石墨阳极.改性石墨阳极具有较小的表面接触角和较高的润湿性,有利于微生物的附着.其电位约为-775 mV,远低于普通石墨阳极的电位(约-450 mV).使用改性石墨阳极的海底生物燃料电池的开路电压约为(1050±50) mV,而普通石墨阳极电池的开路电压为(700±50) mV.改性阳极的海底生物燃料电池输出功率密度可达0.74 W/m2 ,比普通石墨阳极电池高17.4倍.此外,该团队还采用电化学沉积的方法制备了多壁碳纳米管/聚苯胺复合材料修饰的碳毡阳极,并对其电化学性能进行了研究[70 ] .结果表明,改性炭毡阳极的润湿性有所改善,阳极生物膜上的微生物数量比改性前增加了9倍,改性后的海底生物燃料电池的最大功率密度达到0.527 W/m2 ,是改性前的4倍.对阳极材料的改性使制备具有高输出功率密度的海底生物燃料电池成为可能. ...
1
... 一般而言,海底生物燃料电池的输出电压和输出功率较低,限制其广泛应用.因此,目前大多数研究集中于阳极改性上,这是提高电池电压和输出功率的有效手段.Prakash等[68 ] 研究了石墨板和碳布两种阳极材料经表面处理和表面涂层改性后的电性能.该团队制备了4种不同的改性阳极:酸处理碳布、聚苯胺涂覆碳布、酸处理后开槽的石墨板和聚苯胺涂覆普通石墨板.4种阳极均先使用25%的盐酸在40 ℃下加热处理20 min,目的是除去有机成分,同时在缺陷位置引入羧酸基团,有助于在阳极的羧基和苯胺的氨基之间形成氢键.开槽处理目的是在保持阳极尺寸不变的同时增加表面积和表面粗糙度,获得更好的生物膜黏附性,凹槽尺寸为宽1 mm、高0.8 mm.通过实验,得到了不同改性阳极的最大输出功率密度:聚苯胺涂覆碳布为11.14 mW/m2 ,酸处理碳布为5.75 mW/m2 ,聚苯胺涂覆普通石墨板为5.52 mW/m2 ,酸处理后开槽的石墨板为2.86 mW/m2 .研究也证明了聚苯胺涂覆的阳极材料适用于腐蚀性海洋环境,并支持生物膜生长,提高发电量.Fu等[69 ] 报道了一种用电解沉积法制备的铁/氧化铁改性石墨阳极.改性石墨阳极具有较小的表面接触角和较高的润湿性,有利于微生物的附着.其电位约为-775 mV,远低于普通石墨阳极的电位(约-450 mV).使用改性石墨阳极的海底生物燃料电池的开路电压约为(1050±50) mV,而普通石墨阳极电池的开路电压为(700±50) mV.改性阳极的海底生物燃料电池输出功率密度可达0.74 W/m2 ,比普通石墨阳极电池高17.4倍.此外,该团队还采用电化学沉积的方法制备了多壁碳纳米管/聚苯胺复合材料修饰的碳毡阳极,并对其电化学性能进行了研究[70 ] .结果表明,改性炭毡阳极的润湿性有所改善,阳极生物膜上的微生物数量比改性前增加了9倍,改性后的海底生物燃料电池的最大功率密度达到0.527 W/m2 ,是改性前的4倍.对阳极材料的改性使制备具有高输出功率密度的海底生物燃料电池成为可能. ...
1
... 一般而言,海底生物燃料电池的输出电压和输出功率较低,限制其广泛应用.因此,目前大多数研究集中于阳极改性上,这是提高电池电压和输出功率的有效手段.Prakash等[68 ] 研究了石墨板和碳布两种阳极材料经表面处理和表面涂层改性后的电性能.该团队制备了4种不同的改性阳极:酸处理碳布、聚苯胺涂覆碳布、酸处理后开槽的石墨板和聚苯胺涂覆普通石墨板.4种阳极均先使用25%的盐酸在40 ℃下加热处理20 min,目的是除去有机成分,同时在缺陷位置引入羧酸基团,有助于在阳极的羧基和苯胺的氨基之间形成氢键.开槽处理目的是在保持阳极尺寸不变的同时增加表面积和表面粗糙度,获得更好的生物膜黏附性,凹槽尺寸为宽1 mm、高0.8 mm.通过实验,得到了不同改性阳极的最大输出功率密度:聚苯胺涂覆碳布为11.14 mW/m2 ,酸处理碳布为5.75 mW/m2 ,聚苯胺涂覆普通石墨板为5.52 mW/m2 ,酸处理后开槽的石墨板为2.86 mW/m2 .研究也证明了聚苯胺涂覆的阳极材料适用于腐蚀性海洋环境,并支持生物膜生长,提高发电量.Fu等[69 ] 报道了一种用电解沉积法制备的铁/氧化铁改性石墨阳极.改性石墨阳极具有较小的表面接触角和较高的润湿性,有利于微生物的附着.其电位约为-775 mV,远低于普通石墨阳极的电位(约-450 mV).使用改性石墨阳极的海底生物燃料电池的开路电压约为(1050±50) mV,而普通石墨阳极电池的开路电压为(700±50) mV.改性阳极的海底生物燃料电池输出功率密度可达0.74 W/m2 ,比普通石墨阳极电池高17.4倍.此外,该团队还采用电化学沉积的方法制备了多壁碳纳米管/聚苯胺复合材料修饰的碳毡阳极,并对其电化学性能进行了研究[70 ] .结果表明,改性炭毡阳极的润湿性有所改善,阳极生物膜上的微生物数量比改性前增加了9倍,改性后的海底生物燃料电池的最大功率密度达到0.527 W/m2 ,是改性前的4倍.对阳极材料的改性使制备具有高输出功率密度的海底生物燃料电池成为可能. ...
1
... 海底生物燃料电池相较于普通燃料电池具有许多优点.例如,其利用海底沉积层/海水界面作为质子交换膜,不需要人工膜材料,而一般的燃料电池需要昂贵的质子交换膜;阴极自发发生氧的还原反应,不需要催化剂,一般燃料电池的阴极需要贵金属催化剂(如金属铂催化剂);海水和海泥作为电极电解质的一部分,富含盐分、导电性高、内阻小,有利于电池电能输出,满足长期供电需求.植物微生物燃料电池技术是一项全新的低碳环保产能技术,其可以有效利用光能和生物质能等可再生能源来产生电能,具有提供清洁电能的巨大潜力[71 ] .由于微生物可以降解有机物,该技术还可用于污水处理[72 ] .所以该技术应用前景广泛,对于缓解能源危机以及减少环境压力都具有重要的意义,近年来已经成为国际能源领域与环境领域的研究热点.植物微生物燃料电池技术在植物选型之后,需要对植物进行培育,构筑固定式发电系统.对于水稻、大米草等农作物,可以将一定范围内的水稻等进行集中种植,采取统一管理,进行科学生产.这样有利于电极、导线等的设置,构建大规模植物发供电系统,也能帮助农户解决种植中的一些技术和组织管理问题[73 ] .集中式植物供电系统也具备供电成本低的优点.但是未来社会对电力供应的质量与可靠性要求会越来越高,集中式植物供电系统中局部故障所产生的扰动会对整个供电系统造成较大影响,严重时可能引起大面积停电甚至系统的崩溃;集中式系统还易受到战争或恐怖势力的破坏,危害国家用电安全.另外,在不同时间段的用电负荷会发生变化,用电高峰时,电网往往超负荷.集中式大电网不能跟踪电力负荷的变化,调峰能力弱. ...
1
... 海底生物燃料电池相较于普通燃料电池具有许多优点.例如,其利用海底沉积层/海水界面作为质子交换膜,不需要人工膜材料,而一般的燃料电池需要昂贵的质子交换膜;阴极自发发生氧的还原反应,不需要催化剂,一般燃料电池的阴极需要贵金属催化剂(如金属铂催化剂);海水和海泥作为电极电解质的一部分,富含盐分、导电性高、内阻小,有利于电池电能输出,满足长期供电需求.植物微生物燃料电池技术是一项全新的低碳环保产能技术,其可以有效利用光能和生物质能等可再生能源来产生电能,具有提供清洁电能的巨大潜力[71 ] .由于微生物可以降解有机物,该技术还可用于污水处理[72 ] .所以该技术应用前景广泛,对于缓解能源危机以及减少环境压力都具有重要的意义,近年来已经成为国际能源领域与环境领域的研究热点.植物微生物燃料电池技术在植物选型之后,需要对植物进行培育,构筑固定式发电系统.对于水稻、大米草等农作物,可以将一定范围内的水稻等进行集中种植,采取统一管理,进行科学生产.这样有利于电极、导线等的设置,构建大规模植物发供电系统,也能帮助农户解决种植中的一些技术和组织管理问题[73 ] .集中式植物供电系统也具备供电成本低的优点.但是未来社会对电力供应的质量与可靠性要求会越来越高,集中式植物供电系统中局部故障所产生的扰动会对整个供电系统造成较大影响,严重时可能引起大面积停电甚至系统的崩溃;集中式系统还易受到战争或恐怖势力的破坏,危害国家用电安全.另外,在不同时间段的用电负荷会发生变化,用电高峰时,电网往往超负荷.集中式大电网不能跟踪电力负荷的变化,调峰能力弱. ...
1
... 海底生物燃料电池相较于普通燃料电池具有许多优点.例如,其利用海底沉积层/海水界面作为质子交换膜,不需要人工膜材料,而一般的燃料电池需要昂贵的质子交换膜;阴极自发发生氧的还原反应,不需要催化剂,一般燃料电池的阴极需要贵金属催化剂(如金属铂催化剂);海水和海泥作为电极电解质的一部分,富含盐分、导电性高、内阻小,有利于电池电能输出,满足长期供电需求.植物微生物燃料电池技术是一项全新的低碳环保产能技术,其可以有效利用光能和生物质能等可再生能源来产生电能,具有提供清洁电能的巨大潜力[71 ] .由于微生物可以降解有机物,该技术还可用于污水处理[72 ] .所以该技术应用前景广泛,对于缓解能源危机以及减少环境压力都具有重要的意义,近年来已经成为国际能源领域与环境领域的研究热点.植物微生物燃料电池技术在植物选型之后,需要对植物进行培育,构筑固定式发电系统.对于水稻、大米草等农作物,可以将一定范围内的水稻等进行集中种植,采取统一管理,进行科学生产.这样有利于电极、导线等的设置,构建大规模植物发供电系统,也能帮助农户解决种植中的一些技术和组织管理问题[73 ] .集中式植物供电系统也具备供电成本低的优点.但是未来社会对电力供应的质量与可靠性要求会越来越高,集中式植物供电系统中局部故障所产生的扰动会对整个供电系统造成较大影响,严重时可能引起大面积停电甚至系统的崩溃;集中式系统还易受到战争或恐怖势力的破坏,危害国家用电安全.另外,在不同时间段的用电负荷会发生变化,用电高峰时,电网往往超负荷.集中式大电网不能跟踪电力负荷的变化,调峰能力弱. ...
1
... 海底生物燃料电池相较于普通燃料电池具有许多优点.例如,其利用海底沉积层/海水界面作为质子交换膜,不需要人工膜材料,而一般的燃料电池需要昂贵的质子交换膜;阴极自发发生氧的还原反应,不需要催化剂,一般燃料电池的阴极需要贵金属催化剂(如金属铂催化剂);海水和海泥作为电极电解质的一部分,富含盐分、导电性高、内阻小,有利于电池电能输出,满足长期供电需求.植物微生物燃料电池技术是一项全新的低碳环保产能技术,其可以有效利用光能和生物质能等可再生能源来产生电能,具有提供清洁电能的巨大潜力[71 ] .由于微生物可以降解有机物,该技术还可用于污水处理[72 ] .所以该技术应用前景广泛,对于缓解能源危机以及减少环境压力都具有重要的意义,近年来已经成为国际能源领域与环境领域的研究热点.植物微生物燃料电池技术在植物选型之后,需要对植物进行培育,构筑固定式发电系统.对于水稻、大米草等农作物,可以将一定范围内的水稻等进行集中种植,采取统一管理,进行科学生产.这样有利于电极、导线等的设置,构建大规模植物发供电系统,也能帮助农户解决种植中的一些技术和组织管理问题[73 ] .集中式植物供电系统也具备供电成本低的优点.但是未来社会对电力供应的质量与可靠性要求会越来越高,集中式植物供电系统中局部故障所产生的扰动会对整个供电系统造成较大影响,严重时可能引起大面积停电甚至系统的崩溃;集中式系统还易受到战争或恐怖势力的破坏,危害国家用电安全.另外,在不同时间段的用电负荷会发生变化,用电高峰时,电网往往超负荷.集中式大电网不能跟踪电力负荷的变化,调峰能力弱. ...
1
... 除了利用活体植物发电,合理利用农作物秸秆等废弃生物质材料亦可实现高效清洁发电.秸秆、干柴等植物废弃物不完全燃烧时产生的烟气中包含碳氢化合物、一氧化碳、二氧化碳、水蒸气和氮氧化物等物质,将烟气作为固体氧化物燃料电池燃料可以实现清洁供电[74 ] .该技术既获取了电能,又实现了对秸秆等植物废弃物的合理利用,避免了随意燃烧秸秆对环境造成的污染.在军事方面,野外作战不易实现电力补给,利用该途径进行发供电,可以有效解决这一问题,而且野外容易获得植物干柴,燃料来源丰富.此外,基于植物燃料的固体氧化物燃料电池催化裂解植物不完全燃烧产生的烟气中的一氧化碳等小分子物质,减少了黑烟的产生,隐蔽性好,是一项具有发展前景的技术.植物微生物燃料电池发供电技术相比于其他技术来说不仅能产电,还能用于污水处理[75 ] ,而且不需昂贵的材料来构造电池,成本低;系统能够自我修复,生命周期长;该技术以植物作为有机物源,微生物作为催化剂,可广泛用于自然环境中;通过使用混合微生物菌落,可以产生协同作用,提高产电性能,支持高效供电系统的开发. ...
1
... 除了利用活体植物发电,合理利用农作物秸秆等废弃生物质材料亦可实现高效清洁发电.秸秆、干柴等植物废弃物不完全燃烧时产生的烟气中包含碳氢化合物、一氧化碳、二氧化碳、水蒸气和氮氧化物等物质,将烟气作为固体氧化物燃料电池燃料可以实现清洁供电[74 ] .该技术既获取了电能,又实现了对秸秆等植物废弃物的合理利用,避免了随意燃烧秸秆对环境造成的污染.在军事方面,野外作战不易实现电力补给,利用该途径进行发供电,可以有效解决这一问题,而且野外容易获得植物干柴,燃料来源丰富.此外,基于植物燃料的固体氧化物燃料电池催化裂解植物不完全燃烧产生的烟气中的一氧化碳等小分子物质,减少了黑烟的产生,隐蔽性好,是一项具有发展前景的技术.植物微生物燃料电池发供电技术相比于其他技术来说不仅能产电,还能用于污水处理[75 ] ,而且不需昂贵的材料来构造电池,成本低;系统能够自我修复,生命周期长;该技术以植物作为有机物源,微生物作为催化剂,可广泛用于自然环境中;通过使用混合微生物菌落,可以产生协同作用,提高产电性能,支持高效供电系统的开发. ...
1
... 自古至今,人们一直在寻找从自然界中高效获取和利用能量的新方法,自然界中的机械能是可再生的,在地球上处处存在[76 ] .基于电磁机制的传统发电机可以收集大量高频机械能,但分布在环境中的低频歇、随机能量很难被收集和利用[77 ] .植物区域离子浓度扰动发供电技术可以收集和利用自然环境中的低频机械能,并将其转化为电能.其原理为:植物体内的离子浓度在固定区域基本是维持不变的,但是当植物受到外界影响,如风吹扰动、雨滴摩擦等,均会让植物局部的离子浓度发生变化[78 ] .所以,利用压电效应[79 ] 、摩擦电效应[80 ] 等,不仅可以有效地将植物的离子浓度变化变成电能,也能通过植物与自然界中风、雨的振动和摩擦等产生电流,实现能量收集和发供电.国内外一些团队在有关方面开展的工作如下. ...
1
... 自古至今,人们一直在寻找从自然界中高效获取和利用能量的新方法,自然界中的机械能是可再生的,在地球上处处存在[76 ] .基于电磁机制的传统发电机可以收集大量高频机械能,但分布在环境中的低频歇、随机能量很难被收集和利用[77 ] .植物区域离子浓度扰动发供电技术可以收集和利用自然环境中的低频机械能,并将其转化为电能.其原理为:植物体内的离子浓度在固定区域基本是维持不变的,但是当植物受到外界影响,如风吹扰动、雨滴摩擦等,均会让植物局部的离子浓度发生变化[78 ] .所以,利用压电效应[79 ] 、摩擦电效应[80 ] 等,不仅可以有效地将植物的离子浓度变化变成电能,也能通过植物与自然界中风、雨的振动和摩擦等产生电流,实现能量收集和发供电.国内外一些团队在有关方面开展的工作如下. ...
1
... 自古至今,人们一直在寻找从自然界中高效获取和利用能量的新方法,自然界中的机械能是可再生的,在地球上处处存在[76 ] .基于电磁机制的传统发电机可以收集大量高频机械能,但分布在环境中的低频歇、随机能量很难被收集和利用[77 ] .植物区域离子浓度扰动发供电技术可以收集和利用自然环境中的低频机械能,并将其转化为电能.其原理为:植物体内的离子浓度在固定区域基本是维持不变的,但是当植物受到外界影响,如风吹扰动、雨滴摩擦等,均会让植物局部的离子浓度发生变化[78 ] .所以,利用压电效应[79 ] 、摩擦电效应[80 ] 等,不仅可以有效地将植物的离子浓度变化变成电能,也能通过植物与自然界中风、雨的振动和摩擦等产生电流,实现能量收集和发供电.国内外一些团队在有关方面开展的工作如下. ...
1
... 自古至今,人们一直在寻找从自然界中高效获取和利用能量的新方法,自然界中的机械能是可再生的,在地球上处处存在[76 ] .基于电磁机制的传统发电机可以收集大量高频机械能,但分布在环境中的低频歇、随机能量很难被收集和利用[77 ] .植物区域离子浓度扰动发供电技术可以收集和利用自然环境中的低频机械能,并将其转化为电能.其原理为:植物体内的离子浓度在固定区域基本是维持不变的,但是当植物受到外界影响,如风吹扰动、雨滴摩擦等,均会让植物局部的离子浓度发生变化[78 ] .所以,利用压电效应[79 ] 、摩擦电效应[80 ] 等,不仅可以有效地将植物的离子浓度变化变成电能,也能通过植物与自然界中风、雨的振动和摩擦等产生电流,实现能量收集和发供电.国内外一些团队在有关方面开展的工作如下. ...
1
... 自古至今,人们一直在寻找从自然界中高效获取和利用能量的新方法,自然界中的机械能是可再生的,在地球上处处存在[76 ] .基于电磁机制的传统发电机可以收集大量高频机械能,但分布在环境中的低频歇、随机能量很难被收集和利用[77 ] .植物区域离子浓度扰动发供电技术可以收集和利用自然环境中的低频机械能,并将其转化为电能.其原理为:植物体内的离子浓度在固定区域基本是维持不变的,但是当植物受到外界影响,如风吹扰动、雨滴摩擦等,均会让植物局部的离子浓度发生变化[78 ] .所以,利用压电效应[79 ] 、摩擦电效应[80 ] 等,不仅可以有效地将植物的离子浓度变化变成电能,也能通过植物与自然界中风、雨的振动和摩擦等产生电流,实现能量收集和发供电.国内外一些团队在有关方面开展的工作如下. ...
1
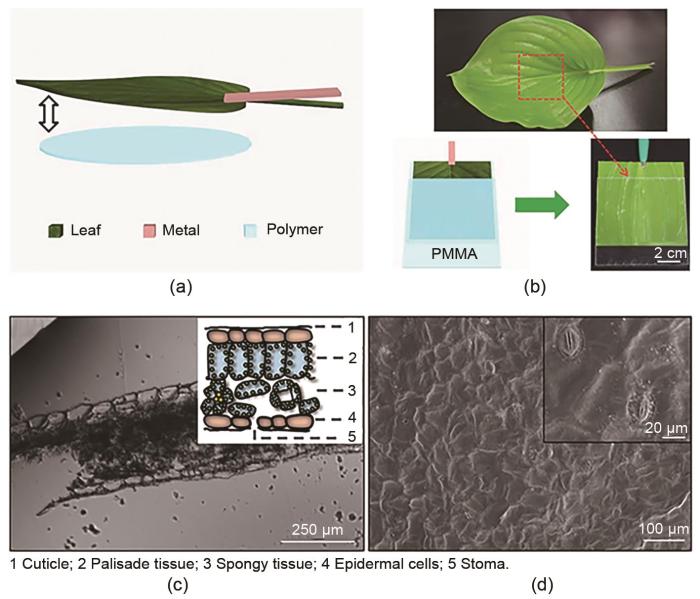
... Chen等[81 ] 开创性地利用植物器官上的摩擦生电效应,使用玫瑰和聚甲基丙烯酸甲酯设计了摩擦电纳米发电机.在频率为2 Hz的恒定周期性压缩力(100 N)下,一片玫瑰花瓣(3 cm×3 cm)产生了30.6 V的最大开路电压和0.78 μA的短路电流.此外,在电阻为50 MΩ的条件下,得到的最大输出功率密度高达27.2 mW/m2 .进一步的研究表明,这些表面富含微纳结构的玫瑰花瓣具有良好的疏水性,意味着其可以产生一定量的电荷并收集风和雨滴的机械能.图17 (a)为实验所使用的天然中国红玫瑰,新鲜玫瑰花瓣从图中黄色区域部分取得.整个发电机的工作机制如图17 (c)所示,底部和顶部的聚甲基丙烯酸甲酯悬臂提供刺激或压力,中间的聚甲基丙烯酸甲酯悬臂的表面通过双面胶粘有花瓣,每个花瓣的底部都有1根铜线作为导体.其末端连接有垫片,制造间距,从而提高振动的效率.当该装置振动1次时,中间板在每个循环中有两次机会接触到顶部和底部的悬臂,这个过程至少可以点亮7个商用LED灯,如图17 (b)所示. ...
1
... Wu等[82 ] 以叶表皮作为电介质和摩擦电材料,导电的叶组织作为底部电极,水滴作为摩擦电材料和顶部电极,设计了1个基于植物叶片和水滴的可完全生物降解的摩擦电纳米发电机,如图18 (a)所示.通过使用这种摩擦电纳米发电机,利用水滴对植物叶片进行扰动,获得了超过1 V和4 μA的电压和电流,如图18 (b)、(c)所示.图18 (d)、(e)分别为水滴撞击叶片表面的示意图和撞击带电的原理图.①滴撞击带电的叶片表面并产生极化,在表面电荷的诱导下,叶/水界面形成双电层.②当液滴扩散到最大面积并接触导线时,液滴充当电极,并将叶片表面与外部电路连接.为了平衡电极之间的电势差,外电路中的电子从植物组织的末端移动到水滴的末端,从而形成电流.③水滴收缩,水/叶接触面积减小.随着双电层面积的减少,电子向后移动到植物组织末端,使电流反向.④液滴离开表面,电流降至0.由于自然界中存在大量的植物叶片和水滴,所以可以利用自然界中水滴对植物的扰动,进行自然能源的采集和植物传感器的研究. ...
1
... Jie等[83 ] 利用玉簪叶设计了一种树叶制成的摩擦电纳米发电机,通过外界扰动获得电能.图19 (a)所示为发电机的结构设计,叶片作为摩擦层和电极,金属电极与树叶接触,构成电路连接,聚合物薄片作为接触层,其与叶子的表面相比电子亲和性差异显著.图19 (b)所示为玉簪叶,其大且亮有光泽.为了研究该发电机的性能,将玉簪叶附着在聚甲基丙烯酸甲酯(具有优异的冲击强度和较轻的重量)上.图19 (c)、(d)分别为玉簪叶的横截面和不规则表面.实验证明,该发电机具有良好的环境适应性,最大输出功率约为45 mW/m2 ,能够驱动一些广告用LED灯和商用电子温度传感器等.这项工作为分布式环境友好型发电机提供了新的发展前景,并在物联网和自供电系统中有应用潜力. ...
1
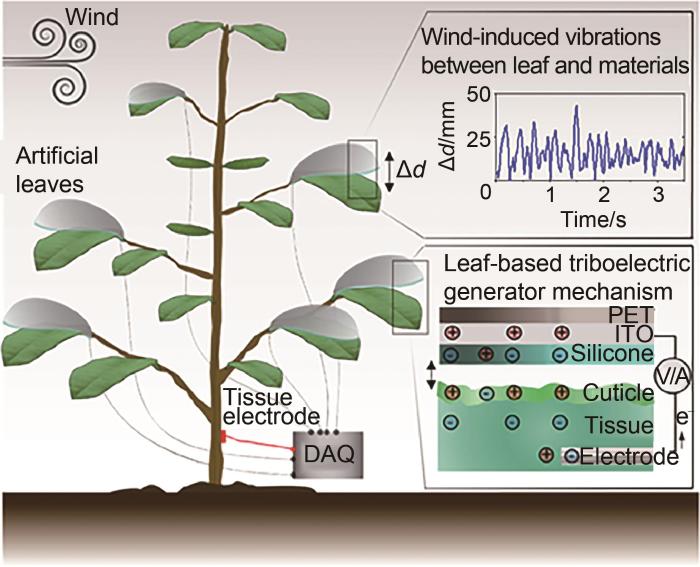
... Meder团队[84 ] 通过分析天然叶片和安装在植物叶柄上的人造硅橡胶叶片之间由风扰动所产生的能量,报告了一种基于活体植物的风能收集系统发电策略.图20 为植物与风的能量收集系统示意图.在风的吹动下,人工硅橡胶叶片与自然叶片一起振动,在叶片之间产生瞬时接触,通过接触带电在植物和人造叶片上产生表面电荷.这些电荷被感应到植物内部组织和人工叶片的电极中,产生电流,从而实现数据采集和能量收集.该团队通过研究无风、有风、有风三种情况下风和人工叶片对输出电压信号的影响,证明在更大风力的扰动下,由于振动幅值、冲击力以及有效接触面积的增大,植物叶片输出电压逐步提高.实验结果表明,1株含有4片人造叶子的夹竹桃可以为多达50个发光二极管和1个带有显示器的数字热传感器供电,证明该能量收集系统可以产生较高的能量转换.其优势还在于可以同时从单个植物的多个叶片收集能量以提高能量输出,且没有因为能量收集过程而对叶片产生伤害. ...
1
... 植物区域离子浓度扰动发供电技术是一种新型的能量收集技术,可以将自然界中的机械能转化为绿色电能,其受地域限制小,从偏远地区到大城市,从森林深处到沙漠,在有植物存在的大多数地区都可以应用.植物机械能的产生是永无止境的过程,可以供长期利用.该技术对植物体生理过程影响也小,因此电池寿命较长.另外,部分装置使用聚合物等作为摩擦材料,可以通过使用废弃聚合物使难以降解的有机污染物得到再利用[85 -86 ] . ...
1
... 植物区域离子浓度扰动发供电技术是一种新型的能量收集技术,可以将自然界中的机械能转化为绿色电能,其受地域限制小,从偏远地区到大城市,从森林深处到沙漠,在有植物存在的大多数地区都可以应用.植物机械能的产生是永无止境的过程,可以供长期利用.该技术对植物体生理过程影响也小,因此电池寿命较长.另外,部分装置使用聚合物等作为摩擦材料,可以通过使用废弃聚合物使难以降解的有机污染物得到再利用[85 -86 ] . ...
1
... 类光合作用发供电技术通过微生物的类光合作用,制备生物光伏电池,为有大片光合微生物存在的固定区域提供电能供应,有望发展新型、可持续的能源供应系统;类光合作用可以吸收二氧化碳,所以该技术在除碳方面具有应用价值.此外,还有一种利用植物光合作用产生的有机物实现发供电的技术,即仿生催化电极(酶催化电极)发供电技术.该技术通过在电极表面修饰生物蛋白酶,借助酶的生物催化活性,结合电化学手段对植物体内的小分子糖等中间产物进行分解,诱导电子的定向移动产生电流.仿生催化电极发供电技术通过将酶修饰的电极嵌入植物体内,与一般的酶燃料电池技术(酶与介体一起溶解在底物中或固定在电极上,利用外加的糖类、甲醇等底物供电)相比,在发电的同时还可实现有机物的原位催化.相较于在植物体外,酶在植物体内更易保持活性,所以电池的长期工作稳定性好;植物的光合作用可以源源不断地提供有机物供酶催化,实现被催化物质的可持续补给;催化得到的产物可以借助于导管和筛管运输,用于植物的生命活动,如呼吸作用等.但该技术在传递质子的过程中,易受到生物膜的阻碍,导致电能转换效率较低.对于一般的酶燃料电池技术,虽然酶在生物体外保持较长时间的催化活性比较困难,使得电池稳定性差,但由于没有传质壁垒,因此会产生更高的电流和输出功率,可以满足一些微型电子设备或生物传感器等对电能的需求[87 ] .仿生催化电极发供电技术受内阻影响较大,如电极的有效面积和电极间距等,质子传输也易受生物膜的阻碍,供电能力弱.此外,酶的成本也较高,活性难控制,使用寿命较其他发供电技术短,还需要考虑酶与植物体的相容性问题.所以该技术构建发供电系统的瓶颈较高,但是其可以发展其他用途,如利用合成生物学原理,依托植物体内的物质和修饰电极的酶等,精准设计合成在传统化学路线上难以合成的物质或药物中间体,这在未来或许比获取电能更有意义. ...
1
... 类光合作用发供电技术通过微生物的类光合作用,制备生物光伏电池,为有大片光合微生物存在的固定区域提供电能供应,有望发展新型、可持续的能源供应系统;类光合作用可以吸收二氧化碳,所以该技术在除碳方面具有应用价值.此外,还有一种利用植物光合作用产生的有机物实现发供电的技术,即仿生催化电极(酶催化电极)发供电技术.该技术通过在电极表面修饰生物蛋白酶,借助酶的生物催化活性,结合电化学手段对植物体内的小分子糖等中间产物进行分解,诱导电子的定向移动产生电流.仿生催化电极发供电技术通过将酶修饰的电极嵌入植物体内,与一般的酶燃料电池技术(酶与介体一起溶解在底物中或固定在电极上,利用外加的糖类、甲醇等底物供电)相比,在发电的同时还可实现有机物的原位催化.相较于在植物体外,酶在植物体内更易保持活性,所以电池的长期工作稳定性好;植物的光合作用可以源源不断地提供有机物供酶催化,实现被催化物质的可持续补给;催化得到的产物可以借助于导管和筛管运输,用于植物的生命活动,如呼吸作用等.但该技术在传递质子的过程中,易受到生物膜的阻碍,导致电能转换效率较低.对于一般的酶燃料电池技术,虽然酶在生物体外保持较长时间的催化活性比较困难,使得电池稳定性差,但由于没有传质壁垒,因此会产生更高的电流和输出功率,可以满足一些微型电子设备或生物传感器等对电能的需求[87 ] .仿生催化电极发供电技术受内阻影响较大,如电极的有效面积和电极间距等,质子传输也易受生物膜的阻碍,供电能力弱.此外,酶的成本也较高,活性难控制,使用寿命较其他发供电技术短,还需要考虑酶与植物体的相容性问题.所以该技术构建发供电系统的瓶颈较高,但是其可以发展其他用途,如利用合成生物学原理,依托植物体内的物质和修饰电极的酶等,精准设计合成在传统化学路线上难以合成的物质或药物中间体,这在未来或许比获取电能更有意义. ...







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}